पुराप्राणिविज्ञान : पूर्वीच्या भूवैज्ञानिक काळातील प्राण्यांसंबंधीचे विज्ञान. हे विज्ञान ⇨पुराजीवविज्ञानाची एक शाखा असून त्याच्यासारखी दुसरी शाखा म्हणजे ⇨ पुरावनस्पतिविज्ञान होय आणि अनुक्रमे प्राण्यांच्या व वनस्पतींच्या जीवाश्मांपासून (शिळारूप झालेल्या अवशेषांपासून) मिळणाऱ्या माहितीवर ही विज्ञाने रचलेली आहेत. त्यांची मूलतत्त्वे, हेतू व उपयोग सारखेच आहेत. प्राण्यांच्या जीवाश्मांचे परीक्षण करून त्यांचे शरीर व राहणी यांविषयी माहिती मिळविणे, त्यांचे वर्गीकरण करणे, त्यांच्या साहाय्याने खडकांची भूवैज्ञानिक वये ठरविणे, खडकांच्या राशीत आढळणाऱ्या जीवाश्मांचा अनुक्रम पाहून प्राण्यांचे निरनिराळे प्रकार कोणत्या क्रमाने पृथ्वीवर अवतरले व त्यांचा कसा विकास झाला यांविषयी माहिती मिळविणेइ. गोष्टींचा समावेश या विज्ञानात होतो [→ जीवाश्म स्तरविज्ञान]. या विज्ञानाच्या विकासासंबंधी इतिहास ‘पुराजीवविज्ञान’ या नोंदीमध्ये दिलेला आहेत.
प्रस्तुत नोंदीत पुराप्राणिविज्ञानाची माहिती प्रथम कालानुक्रमे दिलेली असून नंतर प्राणिगटांनुसार प्रथम अपृष्ठवंशी (पाठीचा कणा नसलेल्या) प्राण्यांची आणि तदनंतर पृष्ठवंशी (पाठीचा कणा असलेल्या) प्राण्यांची माहिती देऊन शेवटी भारतीय संशोधन कार्याची थोडक्यात माहिती दिली आहे. ज्यांच्यात निःसंशय व विपुल जीवाश्म असतात असे सर्वांत जुने खडक म्हणजे कँब्रियन कल्पाच्या (सु. ६० ते ५१ कोटी वर्षांपूर्वीच्या काळाच्या) प्रारंभी व सु. साठ कोटी वर्षांपूर्वी साचलेल्या गाळांचे खडक होत. पुराप्रणिविज्ञानाची ओळख पटण्यासारखे अभिलेख (नोंद) नोंदले जाण्यास तेव्हापासून सुरुवात झाली, असे म्हणण्यास हरकत नाही. त्यापूर्वी काळातील खडकांत कृमींच्या वाटचालीसारख्या खुणा किंवा त्यांनी केलेल्या बिळांसारखी भोके असलेले खडक व स्पंजांच्या कंटिकांसारखे (लहान काट्यांसारखे) आकार असलेले अवशेष इ. प्रकारचे जीवाश्म क्वचित आढळतात पण ते इतके विरळ असतात की, कँब्रियनच्या पूर्वीच्या काळातील खडकांत प्राण्यांचे जीवाश्म जवळजवळ नसतात, असे म्हटले तरी चालेल.
परंतु कँब्रियनच्या आधीच्या कल्पातही प्राणी राहत असले पाहिजेत, एवढेच नव्हे तर ते विपुल व विविध प्रकारचे असले पाहिजे असे मानण्यास पुढील कारणे आहेत : (१) कँब्रियन कल्पाच्या प्रारंभीच्या खडकांत समुद्रात राहणाऱ्या अपृष्ठवंशी प्राण्यांच्या बहुतेक सर्व प्रमुख संघांचे जीवाश्म आढळतात. व ते कित्येक खडकांत विशेष विपुलतेने आढळतात. म्हणजे कँब्रियनच्या प्रारंभीसुद्धा भिन्नभिन्न प्रकारचे विपुल प्राणी राहत असत. त्यांचे पूर्वज त्या काळाच्या पूर्वीही अस्तित्वात असेलच पाहिजेत व ते बरेच विकासही पावले असले पाहिजेत. कँब्रियन कालिन प्राण्यांइतके विकसित व विविध प्राणिप्रकार निर्माण होण्याला दीर्घ कालावधी लागला असला पाहिजे कँब्रियनच्या बऱ्याच आधीच्या (सु. एक अब्ज वर्षांपूर्वीच्या) काळातील समुद्रातही कोणते ना कोणते प्राणी राहत असले पाहिजेत, असा तर्क आहे पण कँब्रियन कालिन प्राण्यांच्या पूर्वजांचे जीवाश्म का सापडू नयेत, हे कोडेच आहे. त्या पूर्वजांना टिकाऊ सांगाडे नसल्यामुळे त्यांचे जीवाश्म होऊ शकले नसावेत. त्यांचा विकास होऊन कँब्रियनच्या सुरुवातीस सांगाडे असणारे प्राणी निर्माण झाले व तेव्हापासून जीवाश्म होण्यास सुरुवात झाली असावी. (२) गाळात पुरल्या गेलेल्या जैव (सेंद्रिय) अवशेषांपासून तयार झालेले ज्या प्रकारचे कार्बन असणारे पदार्थ कँब्रियन कल्पाच्या व त्यानंतर खडकांत आढळतात तशाच प्रकारच्या पदार्थाच्या प्रचंड राशी कँब्रियनच्या पूर्वीच्या खडकांतही आढळतात व त्याही जैव पदार्थांपासून तयार झाल्या असाव्यात.
जीवाश्मांपासून मिळालेल्या माहितीवरून प्राणिकोटीचा जो इतिहास जुळविण्यात आलेला आहे, त्याचे सार सोबतच्या आकृतीस आरेखरूपाने दिलेले आहे. त्याच्यावरून असे कळून येईल की, कवचे असणारे बहुतेक सर्व संघ कँब्रियनच्या सुरुवातीस होते व उरलेले ऑर्डोव्हिसियन कल्पात (सु. ४९ ते ४४ कोटी वर्षांपूर्वीच्या काळात) अवतरले. म्हणजे प्राण्यांचा इतिहास जीवाश्मारूपाने नोंदला जाण्यास सुरुवात झाली तेव्हासुद्धा त्यांचे विविध संघ पृथ्वीवर होते. त्यांचे कित्येक वर्ग व गण ऑर्डोव्हिसियन कल्पात किंवा त्यानंतर अवतरले व पुराजीव महाकल्पाच्या (सु. ६० ते २४. ५ कोटी वर्षांपूर्वीच्या काळाच्या) अखेरीअखेरीपर्यंत त्यांचा विकास व वाढ होत गेली. पुराजीव महाकल्पाच्या अखेरीस पूर्वी प्रमुख असलेले काही गट निर्वंश झाले, पण इतर काहींचा विकास होऊन नवे गट अवतरले व त्यांची वाढ होत गेली. तशाच घटना मध्यजीव महाकल्पात (सु. २३ ते ९ कोटी वर्षांपूर्वीच्या काळात) व त्या महाकल्पांच्या अखेरीस प्राणिकोटीच्या स्वरूपात महत्त्वाचे फेरबदल घडून आले व आधीच्या कित्येक प्राण्यांच्या जागी नवे प्राणी आले. पुराजीव माहाकल्पाच्या उत्तरार्धात जमिनीवर राहणारे प्राणी अवतरले व पृथ्वीवरील प्राण्यांत त्यांचीही भर पडली. पुराजीव महाकल्पाच्या सुरुवातीचे सर्व प्राणी अपृष्ठवंशी होते. पुढे मत्स्य अवतरले. त्या महाकल्पाच्या उत्तरार्धात मत्स्यांची वाढ झाली व प्रथम उभयचर (जमिनीवर पाण्यात राहणारे) व नंतर सरीसृप (सरपटनारे) हे जमिनीवरील प्राणी अवतरले. मध्यजीव महाकल्पात सागरात राहणारे ॲमोनाइट व जमिनीवरील सरीसृप हे प्रमुख प्राणी होते पण त्या महाकल्पाच्या अखेरीस ॲमोनाइट पूर्णपणे व सरीसृप जवळजवळ निर्वंश झाले. मध्यजीव महाकल्पात पक्षी व सस्तन प्राणी अवतरले होते पण ते अतिशय लहान व आदिम (आद्य) प्रकारचे होते. नवजीव महाकल्पात (गेल्या सु. ६.५ कोटी वर्षांत) पृथ्वीतलावर (जमिनीवर) सस्तन प्राण्यांचा आणि पक्ष्यांचा विकास व वाढ झाली. सागरात एकिनॉयडिया, फोरॅमिनीफेरा, गॅस्ट्रोपोडा (शंखधारी) यांची खूप वाढ झाली व ह्याबरोबरच मत्स्य व
बायव्हाल्व्हिया (शिंपाधारी) यांचीही बरीच वाढ होत गेली व प्राणिकोटीला आजच्यासारखे स्वरूप प्राप्त झाले.
केळकर, क. वा.

येथे जीवाश्मांविषयीची माहिती अपृष्ठवंशी व पृष्ठवंशी अशा दोन भागांत दिलेल्या असून अपृष्ठवंशी जीवाश्मांचे प्रमुख आणि गौण गट अशी आखणी विभागणी केली आहे.
अपृष्ठवंशी जीवाश्म-प्रमुख गट : स्ट्रोमॅटोलाइट व अक्रिटार्क : (कँब्रियन-पूर्व म्हणजे सु. ६० कोटी वर्षांच्या आधीचे जीव). जीवाश्म ज्या जीवांचे आहेत दिसते असे त्या जीवांच्या प्रकाराच्या संदर्भात जे निश्चितपणे ओळखू येऊ शकतात व जीववैज्ञानिक वर्गीकरणातील ज्यांचे योग्य स्थान समजू शकते, असे सर्वांतजुने जीवाश्म पुष्कळ देशांमधील कँब्रियन कल्पाच्या अगदी सुरुवातीच्या काळातील खडकांमध्ये आढळले आहेत. तथापि या काळातील ट्रायलोबाइट, स्पंज, ब्रॅकिओपोडा, शैवले इ. जीवांच्या क्रमविकासाची (उत्क्रांतीची) व विभेदनाची (भिन्नभिन्न विशेष प्रकार निर्माण होण्याची) अवस्था इतकी प्रगत झालेली ढळते की, पृथ्वीवर कँब्रियन-पूर्व काळात खूप आधीपासूनच जीव होते. हे मान्य करण्यावाचून गत्यंतर राहत नाही. नंतर या जींवांचा हळूहळू क्रमविकास व विभेदन होत जाऊन कँब्रियन कल्पाच्या अगदी प्रारंभीच्या काळात आढळणाऱ्या विविध गटांच्या अवस्थेपर्यंत ते पोहोचले असले पाहिजेत.
कँब्रियन-पूर्व कालीन खडकांमधील जीवाश्मरूपी अवशेषांचा अभाव लक्षात घेतला, तर त्या खडकांमध्ये विस्तृतपणे आढळणारे सारेच चुनखडक रासायनिक अवक्षेपनाने (साका खाली बसून त्याच्या रूपात साचण्याने) बनले आहेत असा खुलासा करणे अथवा तशी कल्पना करणे शक्य नाही. कँब्रियन-पूर्व कालीन पुष्कळ खडकांच्या स्वरूपाविषयी आपल्याला असलेल्या माहितीवरूनही ते खडक जीवाश्म टिकवून ठेवण्याच्या दृष्टीने योग्य आहेत, असे आपण म्हणू शकतो. काही प्राणिगटांमध्ये सांगाडे अथवा कवच यांच्यासारखे टिकून राहण्यायोग्य असे कठीण भाग (अवयव) निर्माण होऊ शकले नसावेत. हल्लीच्या पुष्कळ प्राण्यांच्या बाबतीतही अशी स्थिती आहे.
कँब्रियन-पूर्व काळातील खडकांमधील जीवाश्मांची उणीव हे १९३० च्या सुमारापर्यंत पुराजीववैज्ञानिकांपुढील एक कोडेच होते. मात्र तेव्हापासून कँब्रियन-पूर्व खडकांमध्ये स्ट्रोमॅटोलाइट नावाचे जीव आढळल्याच्या वार्ता निरनिराळ्या देशांमधून येण्यास सुरुवात झाली. प्राचीन खडकांमध्ये स्ट्रोमॅटोलाइट असल्याचे एकोणिसाव्या शतकातही माहिती होते परंतु तेव्हा त्यांना क्रिप्टोझून म्हणजे संशास्पद जैव अवशेष अथवा आभासी (छद्म) जैव रचना असे संबोधिले जात होते.
स्ट्रोमॅटोलाइट कँब्रियन पूर्व काळाच्या उत्तर (नंतरच्या) भागात मोठ्या प्रमाणात आढळतात, तसेच ते कँब्रियन व ऑर्डोव्हिसियन काळांतही होते परंतु डेव्होनियन (सु. ४० ते ३६.५ कोटी वर्षांपूर्वीच्या काळापासून) त्यांचा जलदपणे ऱ्हास होत जाऊन कार्बॉनिफेरस व पर्मियन (सु. ३५ ते २४.५ कोटी वर्षांपूर्वीच्या) काळांत ते दुर्मिळ झाले.
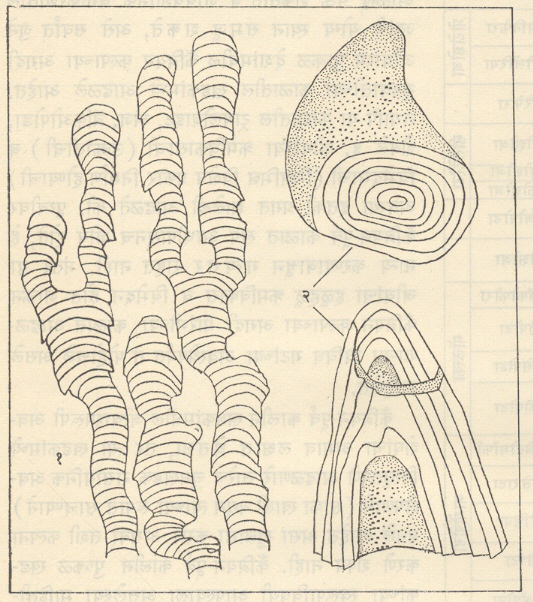
स्ट्रोमॅटोलाइट गुच्छ वा समूहरूपात आढळत असून त्यांचे कोलेनिया, कोनोफायटॉन, बैकालिया, मिंजारिआ इत्यादींसारखे स्थूल संरचनात्माक वर्ग किंवा रूपवंश असे गट करण्यात आले आहेत. स्थूल स्तरवैज्ञानिक सहसंबंध (थरांचे परस्परसंबंध) ठरविणे व कँब्रियन-पूर्व काळातील शैलसमूहांचे (खडकांच्या राशींचे) पोटविभाग पाडणे यांकरिता ते उपयुक्त मानले गेले आहेत.
आताच्या समजुतीनुसार स्ट्रोमॅटोलाइट म्हणजे सामूहिक गटांत राहणाऱ्या विविध नील-हरित शैवलांद्वारे [→ शैवाले] तयार झालेल्या जैव-अवसादी संरचना आहेत. रीसेंट म्हणजे गेल्या सु. ११ हजार वर्षांच्या काळातील स्ट्रोमॅटोलाइट हे बंदिस्त सागरी द्रोणीतील स्वच्छ पाण्यामध्ये आंतरा-वेलीय (भरती-आहोटीच्या) पट्ट्यात १० मी. खोलीपर्यंत आढळतात आणि अधूनमधून ज्यावर पाणी येऊन जाते त्या पंक-मैदानावरही (ओहोटीच्या वेळी उघड्या पडणाऱ्या मंद उताराच्या व चिखलाने आच्छादलेल्या भागावरही) ते आढळतात. अशा प्रकारे स्ट्रोमॅटोलाइटांना ज्याप्रमाणे स्तरवैज्ञानिक सहसंबंधाच्या दृष्टीने महत्त्व आहे त्याचप्रमाणे त्यांच्या हल्लीच्या अधिवासांच्या (राहण्याच्या नैसर्गिक ठिकाणांच्या) दृष्टीने त्यांच्याकडे पाहिल्यास ते ⇨ पुरापरिस्थितिविज्ञानाच्या संदर्भांतही महत्त्वाचे आहेत. स्ट्रोमॅटोलाइट पुष्कळदा फॉस्फेटी निक्षेपांच्या (साठ्यांच्या) समवेत आढळतात, हा त्याच्यासंबंधीचा आर्थिक दृष्ट्या महत्त्वाचा मुद्दा म्हणता येईल.
दक्षिण ऑस्ट्रेलिया, दक्षिण आफ्रिका व इंग्लंड या प्रदेशांतील उत्तर कँब्रियन-पूर्व काळातील निक्षेपांत सीफोझोआ व हायड्रोझोआ यांचे छत्रिक (व्यक्तिगत प्राणी मेड्युसा) म्हणता येतील असे जीवाश्म आढळले आहेत. ॲक्रिटार्क म्हणून ओळखली जाणारी कोशिकीय (पेशीसारखी) स्वरूपाची चिन्हे ही जीवाश्म असल्याचे मानतात. ते विविध प्रदेशांमधील कँब्रियन-पूर्व काळातील खडकांमध्ये आढळल्याचे वृत्त अधूनमधून येत असते. तथापि त्यांच्या उपयुक्ततेची अद्यापि खातरजमा झालेली नाही.
स्ट्रोमॅटोलाइट, सीलेंटेरेटा, ॲनेलिडा व ॲक्रिटार्क यांच्या साहाय्याने कँब्रियन-पूर्व काळातील जीवांविषयीच्या आपल्या माहितीत भर पडते. त्याचप्रमाणे त्या खडकांमध्ये आढळणारे मार्ग-चिन्ह (वाटचालीच्या खुणांच्या स्वरूपाचे पथ-चिन्ह) जीवाश्म त्या काळातील जीवांविषयीच्या माहितीत विशेषकरून ते ज्या शैलसमूहांत आढळतात ते शैलसमूह कोणत्या पर्यावरणीय स्थितीत बहुतकरून निक्षेपित झाले (साचले) असावेत, या विषयीच्या माहितीत भर घालण्याच्या दृष्टीने उपयुक्त ठरतील.
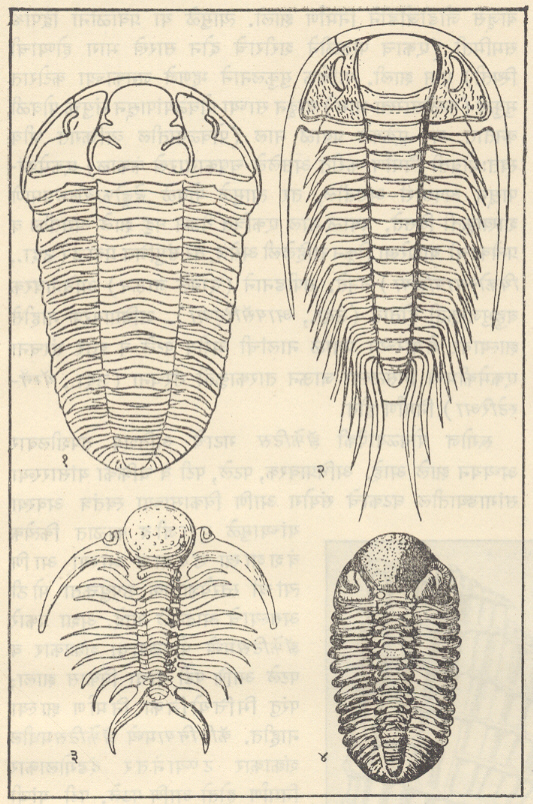
ट्रायलोबिटा : सर्वांत आधी अवतरलेले आर्थ्रोपोडा आणि जीवाश्मांच्या रूपात सर्वांत चांगल्या रीतीने माहीत असलेल्या कवचधारींच्या (क्रस्टेशियनांच्या) दोन गटांपैकी एक गट म्हणजे ट्रायलोबाइटी प्राण्यांचा गट होय. ट्रायलोबाइटांचे जीवाश्म फक्त सागरी निक्षेपांतच आढळले आहेत. ट्रायलोबाइटांच्या बाह्यकंकालाचे (आधार वा संरक्षण देणार्याप बाह्य कठीण संरचनेचे) पुढील तीन प्रमुख भाग असतात : (१) अग्रभागी फलकासारखे शीर्ष व त्याच्या मध्यभागी भ्रूमध्य, (२) बरगडीसारखे कायखंड असलेले वक्ष आणि (३) ज्यावर सामान्यपणे कायखंड दिसतात ते अवसानक [→ ट्रायलोबाइट]. बाह्य कवचावरील दोन उभ्या सीतांमुळे (खोबणींमुळे) पूर्ण शरीर तीन खडांचे बनल्यासारखे दिसते. ट्रायलोबाइटांमध्ये सर्वसामान्यपणे पुढील संरचनात्मक परिवर्तने झालेली आढळतात : शीर्ष व अवसानक यांचे आकारमान वाढणे, उदा., ॲग्नॉस्टस भ्रूमध्य अग्र बाजूस रुंद होणे व भ्रूमध्यीय सीता नाहीशी होणे, उदा., फॅकॉप्स शीर्षावरील कपोलकोन व कायखंडांची टोक मागील बाजूस कंटकरूप होणे, उदा., पॅराडॉक्साइड व कायखंडांची संख्या २९ पासून (उदा., हारपीज) २ पर्यंत (उदा., ॲग्नॉस्टस) घटणे. बऱ्या, चदा हे आकारवैज्ञानिक (प्राण्याचा आकार व संरचना यांच्या दृष्टीने होणारे) बदल एकमेकांशी निगडित नसत. या बदलांच्या संयोगांतून ट्रायलोबाइटांच्या एकानंतर दुसरी अशा अनेक वंश-शाखा उत्पन्न होत गेल्या. या प्रत्येक शाखेचा कालावधी बहुधा मर्यादित असून ट्रायलोबाइटांचा एक गट म्हणून विचार केला, तर त्यांच्या क्रमविकासाचा ओघ अतिशय जटिल असल्याचे दिसते. ट्रायलोबाइटांचे एकेकटे वंश किंवा वंशांचे गट विविध थरांमध्ये विशिष्ट समुच्चयाने आढळत असल्याने थरांच्या मालिकेची ठळक स्तरवैज्ञानिक एककांत (घटकांत) विभागणी करण्यासाठी त्यांची मदत झाली. ट्रायलोबाइट हे निश्चितपणे चलनक्षम प्राणी असून त्यांचे विस्तृत भागांत स्थलांतर झाले होते. अशा प्रकारे दूरवर उघड्या पडलेल्या शैलसमूहांचे सहसंबंध ठरविण्यासाठी ट्रायलोबाइटांचे जीवाश्म उपयुक्त आहेत. ट्रायलोबाइट मोठ्या प्रमाणावर नितलस्थ (सागराच्या तळावर राहणारे) व इतस्ततः रांगणारे असून काही ट्रायलोबाइट सागरी तळाजवळ किंवा उघड्या समुद्रात पोहू लागले होते, असा निष्कर्ष पुढील गोष्टींवरून काढता येतो : पोहण्याच्या दृष्टीने योग्य (अनुकूल) शरीराकृती, संतुलनाचे अवयव म्हणून असलेल्या फुगीर कंटिका, तरंगण्यास उपयुक्त असा मोठा व फुग्यासारखा भ्रमध्य आणि रेमोप्ल्युराइड, इंग्लिना, डायकॉन इ. वंशांमध्ये आढळणारी जटिल व अग्रभागी असलेली दृष्टीअंगे. ॲग्नास्टस, मायक्रोडिस्कस इत्यादींसारख्या काही वंशांतील ट्रॉयलोबाइट तळावरील गाळात खणण्याच्या व गाळ उकरून त्याच्या खालून हिंडण्याच्या सवयीमुळे आंधळे बनले होते, तर डाल्मॅँनाइट व ओलेनेलसारख्या काहींच्या शेपटीवर लांब काटा होता व बहुतकरून तो हल्लीच्या लिम्युलसप्रमाणे तळावरील गाळात स्वतःला खिळवून ठेवण्याकरिता व अन्न मिळवण्याच्या उद्देशाने आपल्या डोक्याने गाळ खणून उपसण्यासाठी ते वापरत असावेत. सिल्युरियन अखेरीपर्यंत (सु. ४२ कोटी वर्षांपूर्वीपर्यंत) ट्रायलोबाइटांची अतिशय भरभराट झाली, तदनंतर डेव्होनियनमध्ये त्यांचा जलदपणे ऱ्हायस झाला व पर्मियनअखेरीस (सु. २७ ते २४.५ कोटी वर्षांपूर्वीच्या काळाच्या शेवटी) किंवा कदाचित ट्रायासिक (सु. २३ ते २० कोटी वर्षांपूर्वीच्या)काळाच्या सुरुवातीलाच कोणतीही वंशशाखा मागे न ठेवता ते निर्वंश झाले.
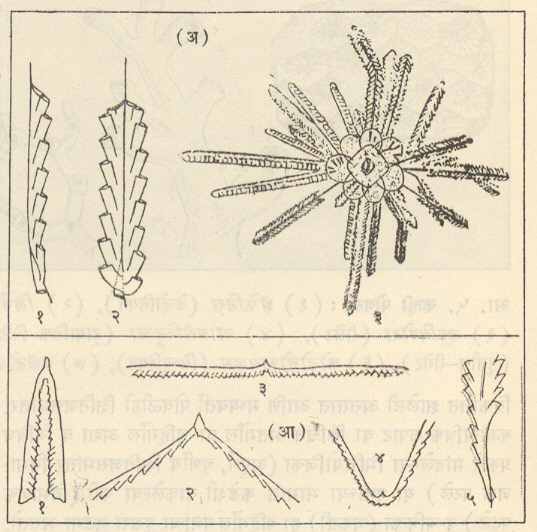
हेमिकॉर्डेटा-ग्रॅप्टोलिथिना : ग्रॅप्टोलाइट हे प्राणी कँब्रियन काळाच्या अखेरीस पृथ्वीवर अवतरले व सिल्युरियन काळाच्या शेवटी निर्वंश झाले [→ ग्रॅप्टोलाइट]. ते केवळ सागरी पर्यावरणातच आढळले आहेत. निर्वंश झालेल्या या प्राणिगटाच्या जीवविज्ञानातील स्थानाविषयी अल्पशीच माहिती आहे.
मात्र कोझलोव्हस्की यांच्या संशोधनानुसार टेरोब्रॅंकच्या ऱ्हाब्डोप्ल्यूरा वंश हा ग्रॅप्टोलाइटांना सर्वांत जवळचा प्राणिगट आहे. ग्रॅप्टोलाइटांचे जीवाश्म सामान्यपणे शेल, शेली चुनखडक व सूक्ष्मकणी वालुकामय शेल या खडकांत आढळतात. हे जीवाश्म म्हणजे सूक्ष्म प्रावार वा कप्पे असलेले बारीक कार्बनीभूत दांडे (वृंत) असून त्यांना रॅब्डोसोम म्हणतात. ते ग्रॅप्टोलाइटच्या पॉलिपमध्ये (व्यक्तिगत जीवामध्ये) क्रमाने मांडलेले असतात व टेट्राग्रॅप्टसामधल्याप्रमाणे मध्य तबकडीला जोडलेले दांडे (रॅब्डोसोम) लोलकाप्रमाणे लोंबते असतात. ग्रॅप्टोलाइटांचा क्रमविकास प्रामुख्याने दोन दिशांनी झाला : (१) दांड्यांची संख्या वाढत्या प्रमाणात ३२ पासून (उदा. क्लोनोग्रप्टस) एकापर्यंत (उदा., मोनोग्रॅप्टस) घटली आणि (२) रॅब्डोसोमांची ठेवण अधिकाधिक अपसारी होत जाऊन ती लोंबती होती त्याऐवजी आरोही किंवा उत्पन्न होत गेली [आ. ४ (आ)]. ज्यांच्या क्रमविकासात कमीअधिक प्रमाणात या दोन्ही दिशांनी सुसूत्र असे बदल झालेत ते ग्रॅप्टोलाइटांचे गट आसमंतातील परिस्थितीशी अधिक जुळवून घेणारे व प्रगतिशील असल्याचे दिसून आले. उलट ज्यांच्या क्रमविकासात वरीलपैकी एका दिशेतील बदल अधिक प्रमाणात झाले होते त्या वंशशाखांची अखेर लवकर झाली व त्यांच्यापासून अगदी थोडेच भिन्न (परिवर्तित) प्रकार निर्माण झाले. वरील दोन दिशांतील सुसूत्र परिवर्तनाच्या जोडीने प्रावरभित्तींची संरचना हा परिवर्तनांमध्ये भर घालणारा तिसरा घटक होता. ग्रॅप्टोलाइट हे सवयीने उघड्या समुद्रात तरंगणारे व महासागरांतील प्रवाहांनुसार वाहत जाणारे गुच्छरूपातील निवह (समूहरूपी वसाहती) होते, असे त्यांच्यातील प्रावरभित्तींच्या संरचनांवरून सूचित होते. त्यांच्या अल्पायुष्याच्या इतक्या थोड्या कालावधीत त्यांचा एवढा विस्तृत प्रसार (फैलाव) कसा झाला, हे यावरून स्पष्ट होते. जलदपणे क्रमविकास होत जाऊन एकापाठोपाठ निर्माण झालेले प्रकार ऑर्डोव्हिसियन व सिल्युरियन कालीन थरांचे स्तरवैज्ञानिक उपविभाग पाडण्यास साहाय्यभूत ठरले, कारण प्रत्येकातील ग्रॅप्टोलाइटांचे जीवाश्म-समूह वैशिष्ट्यपूर्ण आहेत. ग्रॅप्टोलाइटांच्या या स्तरवैज्ञानिक उपयुक्ततेशिवाय त्यांच्या विस्तृत प्रसारामुळे दूरवर आढळणाऱ्या (उदा., स्कँडिनेव्हिया-स्कॉटलंड ते कॅनडापर्यंतचे ग्रॅप्टोलाइटयुक्त थर) शैलसमूहांचे सहसंबंध निश्चित करण्यास त्यांची मदत झाली आहे. ग्रॅप्टोलाइटांच्या जीवाश्मांमुळे पूर्वी ज्याला पूर्व सिल्युरियन (संघ) म्हणत असत त्याला ऑर्डोव्हिसियन संघ असा स्तरविज्ञानातील स्वतंत्र दर्जा प्राप्त झाला, कारण त्याच्यातील ग्रॅप्टोलाइट जीवाश्म हे मूळच्या उत्तर सिल्युरियन संघातील जीवाश्मांच्या मानाने अतिशय वेगळे आहेत. कँब्रियनच्या शेवटी व ऑर्डोव्हिसियन काळाच्या सुरुवातीस दांड्यांची संख्या कमी होण्याची व ते अपसारी होण्याची प्रवृत्ती ह्या ग्रॅप्टोलाइटांच्या क्रमविकासातील सर्वसाधारण पद्धती होत्या. उत्तर ऑर्डोव्हिसियन काळात दोन व चार शाखांचे आणि सिल्युरियन काळात दोन व एक शाखा असलेले वंश सामान्यपणे आढळत होते.
प्रावारांच्या एक श्रेणी व द्विश्रेणी मांडणीची उपपत्ती अद्यापि चांगली कळलेली नाही, म्हणून ग्रॅप्टोलाइटांच्या वंशामधील हल्ली उल्लेखिले जाणारे जातिविकासाचे संबंध स्पष्ट झालेले नाहीत.
मेल्यानंतर ग्रॅप्टोलाइटांचे निवह बुडून समुद्रतळावर गेले. तेथे ते पुष्कळदा कार्बनयुक्त व पायराइटयुक्त काळ्या रंगाच्या खडकांत जोडीने आढळतात. त्यावरून तेथे अनॉक्सिजीवी (ऑक्सिजनरहित अवस्थेत जगण्याची) परिस्थिती सर्वसाधारणपणे अस्तित्वात होती, असे सूचित होते. ग्रॅप्टोलाइट प्राणिजातीच्या प्रसाराच्या दिशांचा काळजीपूर्वक अभ्यास केला, तर त्या प्राचीन काळातील महासागरी प्रवाहांच्या मार्गांचे नकाशे तयार करण्यास उपयुक्त ठरू शकतील. अशा तऱ्हेपने स्तरवैज्ञानिक उपयुक्ततेशिवाय ⇨ पुराभगोल आणि ⇨ पुरापरिस्थितिवाज्ञान यांच्या दृष्टीने ग्रेप्टेलाइट फार महत्त्वाचे आहेत. [→ ग्रॅप्टोलाइट].
सीलेंटेरेटा : (अँथोझोआ-झोअँथेरिया, पोवळे वा प्रवाळ). साध्या आधुनिक झोअँथेरियन पोवळ्याच्या [मॅड्रिपोरॅयन वा स्क्लेरॅरियन वा स्क्लेरॅक्टिनियन → पोवळे] चूर्णीय (कॅल्शियम कार्बोनेटयुक्त कॅल्केरिअस) संरचनेमध्ये बाह्य भित्ती वा अधिप्रावरक असते. त्या भित्तीची उदग्र दिशेत अरीय पद्धतीची सहा आद्य पटले मांडलेली असून ती सामान्यतः मध्यवर्ती कीलापर्यंत (स्तंभिकेपर्यंत) गेलेली असतात. त्यांच्यानंतर अधिकाधिक आखूड द्वितीयक, तृतीयक इ. पटले सहाच्या पटीत विकसित झालेली असतात आणि मध्यवर्ती पोवळीही क्षितिजसमांतर, कमीअधिक सपाट वा किंचित अंतर्गोल वा बहिर्गोल अशा व विविध प्रकारे मांडलेल्या भित्तीयोजिका (अपूर्ण, चूर्णीय क्षितिजसमांतर विभाजक पटले) या पत्रांच्या भागात कडेशी वाकलेल्या पटी (विभाजक पटले) व चक्रिका (चकती) हा बहिर्गोल पत्रांचा दुसरा जुडगा असतो. या सर्वांमुळे पुटिकामय संरचना निर्माण होते. सहा वा सहाच्या पटीत पटले असलेल्या पोवळ्यांच्या या गटाला हेक्झॅकोरॅलिया असेही म्हणतात.
पुराजीवकालीन रूगोज (वलिवत) पोवळ्यांत मुख्य व विरोधी पटले विकसित झाल्यावर नंतरची पटाले दोन आद्य पटलांच्या दोन्ही बाजूंस जोडीजोडीने निर्माण झाली. त्यामुळे या प्रवाळांना द्विपार्थ सममिती (एकाच पातळीने शरीराचे दोन सारखे भाग होण्याची स्थिती) प्राप्त झाली. पार्श्विक मुकुलनाने म्हणजे जनकाच्या कटोरात मुकुल (कळीसारखा भाग) वाढून साध्या पोवळ्यांपासून संयुक्त पोवळी बनतात. जर एकेकटे प्रवाळ नाल (पोवळ्यातील व्यक्तिगत जीव स्वतःभोवती निर्माण करीत असलेले चषकासारखे कंकाल) एकमेकांपासून अपसारी असतील, तर त्यामुळे पोवळे डेंड्रोफायलमप्रमाणे शाखाकृती दिसते. प्रवाळ नाल एकत्रित येऊन घट्ट झाले असतील व प्रत्येकाची बाह्यरेखा टिकून राहिलेली असेल, तर संपुंजित प्रवाळ (उदा., थिकोस्मायलिया) बनते. संपीडनाने (दाबले जाऊन) अधिप्रावरक बहुभुजाकृती होतात (उदा., आयसॅट्रिआ). अधिप्रावरक नाहीसे झाल्यावर शेजारच्या प्रवाळ नालांची अरीय पटले व इतर संरचना एकमेकींमध्ये मिसळल्या जाऊन ताराकृती संरचना (उदा., थॅम्नॅस्टेरिआ) निर्माण होते.
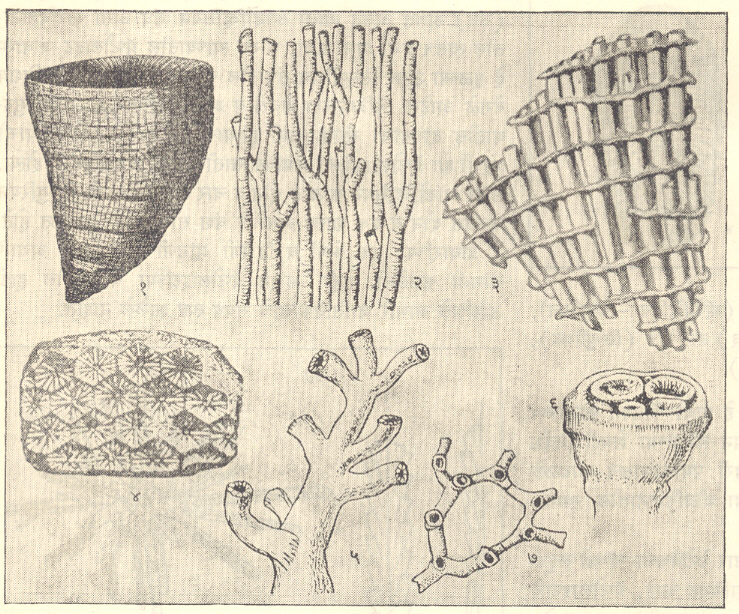
रूगोज पोवळ्यांपैकी झॅफ्रेंटिस गटाचे अतिशय तपशीलवार अध्ययन झाले आहे. अधिप्रावरक, पटले, पटी व चक्रिका यांसारख्या सांगाड्यातील घटकांचे संयोग आणि विकासाच्या स्वतंत्र अवस्था यांच्यामुळे पुराजीव काळात कित्येक वंशशाखा उदयास आल्या आणि त्यांची स्तरवैज्ञानिक उपयुक्तता मोठी असल्याचे आढळले आहे. अशा प्रकारे झॅफ्रेंटिसमध्ये पोवळ्याचा शंक्वाकार व पटले आणि पटी यांचा विकास झाला परंतु भित्तियोजिका निर्माण झाल्या नाहीत. कॅनिनियामध्ये झॅफ्रेंटिसमधील शंक्वाकार टप्प्यांनंतर दंडगोलाकार निर्माण होतो आणि पटले, पटी यांची व भित्तीयोचिकांचीही चांगली वाढ होते. ऑलोफायलममध्ये झॅफ्रेंटिसकॅनिनिया टप्प्यानंतर कडेवरील पट्ट्यांत बहिर्गोल चक्रिकांचा विकास होतो. पॅलिओस्मायलियामध्ये पटी अतिशय बर्हिगोल होतात व त्या चक्रिकेत मिसळून जाण्याची प्रवृत्ती आढळते. लिथोस्ट्रॉशियनमध्ये कील विकसित झालेला असतो परंतु पटी वा चक्रिका अल्पच विकसित झालेल्या असतात. या व सिस्टिफायलम, तसेच इतर संजातीमधील (वंशशाखांमधील) विकासाच्या इतर दिशांवरून रूगोज पोवळ्यांना पुराजीव स्तरविज्ञानात अतिशय महत्त्वाचे स्थान आहे. पुराजीव काळातील रूगोजांप्रमाणे सांगाड्यातील घटकांच्या संयोगांना अनुसरून मॅड्रेपोरॅरियांचा विकास कित्येक दिशांनी झाला आहे.
हेटरोकोरॅलिया हा झोअँथेरियन पोवळ्यांचा एक लहान गट असून तो केवळ कार्बॉनिफेरस (सु.३५ ते ३१ कोटी वर्षांपूर्वीच्या ) काळात आढळतो आणि त्यातील पोवळ्यांची पटले रूगोजप्रमाणे अरीय मांडणीची असतात. त्यांच्यात अधिप्रावरक नसतात परंतु पटी अतिशय बहिर्गोल असून कडेच्या भागात त्या इतक्या उतरत्या असतात की, त्यामुळे लागोपाठच्या पटींच्या कडा एकमेकींत मिसळून अधिप्रावरकाच्या जागी पातळ बाह्यभित्ती तयार झालेली असते.
केवळ पुराजीव काळात आढळणाऱ्या पोवळ्यांमधील सुटे प्रवाळ नाल हे साधे अरुंद नळीसारख्या संरचना असून त्यांना खालील दिशेत असणाऱ्या नसराळ्यासारख्या पटी व झोअँथेरियन पोवळ्यांतील पटलांच्या ऐवजी अनुदैर्घ्य कटक(लांबीच्या दिशेने असलेले उंचवटे) असतात. पार्श्वी दिशेत प्रवाळ नाल क्षितिजसमांतर तटकांनी जोडलेले असतात. पुराजीव महाकल्पातील प्रवाळभित्ती मुख्यत्वे पटीयुक्त प्रवाळांच्या बनलेल्या आहेत.
पटीयुक्त प्रवाळांची संरचना पुष्कळशी ॲल्शिओनेरियाच्या (किंवा ऑक्टोकोरॅलियाच्या) संरचनेसारखी आहे. तथापि ॲल्शिओनेरियाचे जावाश्म विरळाच आढळतात व त्याची उपयुक्तताही थोडीच आहे. पटीयुक्त पोवळ्यांचे रूगोज व मॅड्रेपोरॅरिया यांच्याशी असलेले संबंधही अनिश्चित प्रकारचे आहेत.
झोअँथेरिया ऑर्डोव्हिसियन काळात अवतरल्याचा निश्चित पुरावा उपलब्ध आहे, तर रूगोज पुराजीव काळाच्या शेवटी निर्वंश झाले. मॅड्रेपोरॅरियांच्या ट्रायासिक काळातील नोंदी गोंधळात टाकणाऱ्या आहेत आणि रूगोज व मॅड्रेपोरॅरिया यांच्यातील संबंध अनिश्चित स्वरूपाचे आहेत. पुराजीवाची अखेर ते मध्यजीवाचा प्रारंभ या संक्रमण काळात रूगोजांच्या विविध शाखांपासून मॅड्रेपोरॅरिया उत्पन्न झाले असण्याची शक्यता आहे.
हल्ली एकेकटे मॅड्रेपोरॅरियन पोवळे बहुतेक सर्व अक्षांशांमध्ये व निरनिराळ्या खोलीच्या पट्ट्यांत आढळते. बहुसंख्य १०० ते २,००० मी. खोलीच्या पट्ट्यात व काही त्याच्या पलीकडे आढळतात. खोल सागरात राहणाऱ्या प्रकारांच्या संरचना नाजूक व भंगुर असून त्यांच्यापैकी थोडेच नवजीव काला इतक्या आधीच्या व त्यापेक्षाही थोडे मध्यजीव काळात आढळतात.
सध्याची भित्ती निर्मिणारी पोवळी सागरी आहेत (याला थोडेच अपवाद आहेत). ती सागराचा किनारा ते सु. ३० मी. खोलीपर्यंतच्या मर्यासदित भागात व काही ५० मी. खोलीपर्यंत आढळतात. या भागातील तापमान २५० ते ३०० से. असून पाणी स्वच्छ असते, म्हणून आधुनिक प्रवाळभित्ती अटलांटिकचा उष्ण कटिबंधीय भाग, हिंदी व पॅसिफिक महासागरांचा मध्य व पश्चिम भाग आणि अरबी समुद्र या भागांतच आहेत. उष्ण प्रवाहांच्या परिणामामुळे उष्ण कटिबंधीय भागाच्या बाहेरही त्या तयार झालेल्या आढळतात उदा., बहामा, फ्लॉरिडा व दक्षिण जपान येथील प्रवाळभित्ती. भित्तीनिर्मिणारी आधुनिक पोवळी ज्या मर्यायदांमध्ये राहत आहेत, त्या मर्यादांना प्राचीन काळातील प्रवाळभित्तींच्या बाबतीत पुराभूगोलीय व पुरापरिस्थितिविज्ञानीय दृष्टीने फार महत्त्व आहे. [→ पोवळे].
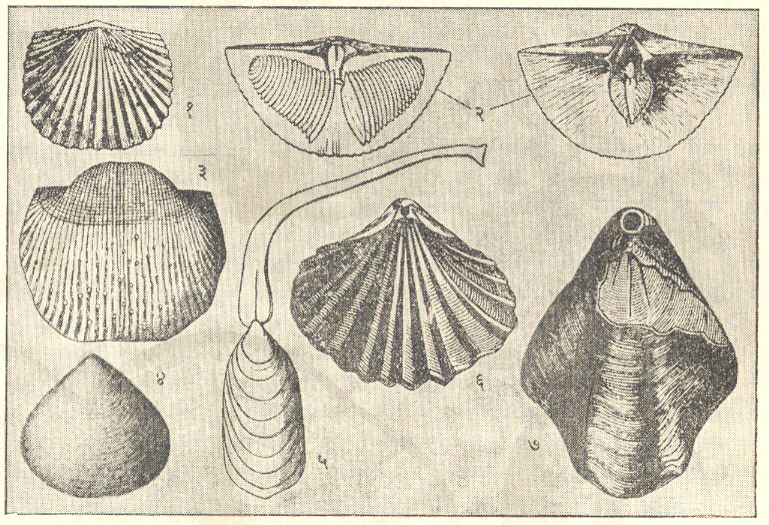
ब्रॅकिओपोडा : सर्व ब्रॅकिओपोडा सागरात राहणारे आहेत. त्यांचे कवच पृष्ठीन व मुखीन अशा दोन कपाटांचे (शकलांचे) बनलेले असून त्यांपैकी मुखीन कपाट नेहमी थोडे मोठे असते. ब्रॅकिओपोडाच्या इनार्टिक्युलेटा या वर्गामध्ये दोन्ही कपाटे पश्च कडेवर केवळ स्नायूंनी तर आर्टिक्युलेटा या वर्गामध्ये मुखीन कपाटातील दोन दात व पुष्ठीन कपाटावरील दोन खोबणी (उखळी), यांच्या बनलेल्या बिजागरीने जोडलेली असतात. या दोन कपाटांतून बाहेर आलेल्या वृंताने (देठाने) प्राणी तळावरील खडकाला चिकटलेला असतो. वृंत ज्यातून बाहेर पडतो त्या छिद्रामध्ये क्रमविकास होताना परिवर्तने झाली आणि या परिवर्तनांच्या आधारावर इनार्टिक्युलेटाचे अट्रेमाटा व निओट्रेमाटा आणि आर्टिक्युलेटाचे पॅलिओट्रेमाटा, प्रोट्रेमाटा व टिलोट्रेमाटा असे पद्धतशीर विभाग केले जातात. आर्टिक्युलेचटामधील स्पिरिफेरॅसिया व टेरेब्रॅटयुलॅसिया यांच्यात बाहूंना आधार देण्याकरिता वेटोळेयुक्त चूर्णीय बाहु-कंकाल म्हणजे स्पायरॅलिया निर्माण झाले होते. ब्रॅकिओपोडांच्या जीवाश्मरूप कवचांतील या सर्व आकारवैज्ञानिक वैशिष्ट्यांचे कपाटांच्या विकासांची पद्धती, कपाटाच्या आतील बाजूवर उमटलेल्या स्नायूंच्या चिन्हांची तऱ्हा व त्यांच्या सूक्ष्मरचना यांच्यानुसार अध्ययन करणे शक्य असते व या अध्ययनाची या गटांचा जातीविकास व क्रमविकासाचा ओघ यांचा मागोवा घेण्यास मदत होते. ब्रॅकिओपोडा प्राणी जरी खडकाला चिकटून राहणारे असले, तरी डिंभावस्थेच्या (भ्रूणानंतरच्या स्वतंत्रपणे अन्न मिळवून जगणाऱ्या आणि प्रौढाशी साम्य नसणाऱ्या सामान्यतः क्रियाशील पूर्व अवस्थेच्या) दीर्घ काळामध्ये त्यांचा प्रसार झाला असला पाहिजे व त्यामुळे त्यांचा प्रसार विस्तृत झालेला दिसतो.
जरी विविधता व संख्या यांच्या दृष्टीने ब्रॅकिओपोडा पूर्ण पुराजीव काळात हळूहळू विपुल होत गेलेले दिसत असले, तरी काही स्थानिक व अगदी लहान प्रदेशांचा अपवाद वगळता ते सामान्यपणे स्तरवैज्ञानिक सहसंबंध ठरविण्याकरिता उपयुक्त ठरले नाहीत. तथापि स्पिरिफर सिरिंगोथायरिस व प्रॉटक्टस आणि त्यांच्याशी संबंधित असलेल्या ब्रॅकिओपोडांचा विशेषतः टेथिस समुद्रातील आल्प्स-हिमालय पट्ट्यात कार्बॉनिफेरस व पर्मियन काळात फार विकास व प्रसार झाला होता आणि तेथील थरांचे सहसंबंध ठरविण्यासाठी ते उपयुक्त आहेत. ज्यांची पुराजीव काळात भरभराट झाली असे बहुतेक ब्रॅकिओपोडा प्राणी पर्मियन काळाअखेर निर्वंश झाले. त्यांपैकी काही थोड्याच वंशशाखा मध्यजीव काळात टिकून राहिल्या होत्या व टिलोट्रेमाटा हा त्यांपैकी प्रमुख गट होय. अगदी थोडे स्फिरिफर गटाचे काही वंशज ट्रायासिक काळात जगण्याची धडपड करीत होते. जुरासिक (सु. १८. ५ ते १५.५ कोटी वर्षांपूर्वीच्या) व क्रिटेशस (सु. १४ ते ९ कोटी वर्षांपूर्वीच्या) काळात टेरेब्रॅट्युलॅसिया व ऱ्हिंकोनेलॅसिया हे प्रमुख गट होते. नवजीव महाकल्पात ब्रॅकिओपोडांचा र्हास जलद झाला व हल्ली त्यांचे जास्तीत जास्त ६० वंश असून त्यांपैकी बहुसंख्य आर्टिक्युलेटा गटातील आहेत. लिंग्यूला, क्रेनिया व डिसिना हे इनार्टिक्युलेटाचे वंश हल्ली आढळत असून ते ऑर्डोव्हिसियन काळापासून आतापर्यंत इतका आश्चर्यकारक दीर्घकाळ टिकून राहिले आहेत.
ब्रॅकिओपोडा आढळण्याचे प्रदेश सामान्यतः मॉलस्कांच्या (मृदुकाय प्राण्यांच्या) प्रदेशांशी निगडित असतात. बहुसंख्य इनार्टिक्युलाटा ओहोटीच्या पातळीपासून २० मी. खोलीपर्यंतच्या पट्ट्यात व उरलेले थोडेच १०० मी. खोलीपर्यंत आढळतात. आर्टिक्युलेटा सामान्यपणे १५ ते १०० मी. खोलीच्या पट्ट्यात तर टेरेब्रॅट्युला गटाचे प्राणी किनाऱ्यापासून १०० मी. खोलीपर्यंत आढळतात. ऱ्हिंकोनेलिड गटाचे प्राणी सर्व अक्षांशांत आढळतात पण बहुसंख्यांना थंड पाणी अधिक पसंत असून ते १५ ते १०० मी. खोलीच्या पट्ट्यात आढळतात. अशा प्रकारे गॅस्ट्रोपोडा आणि लॅमेलिब्रँकिया (शिंपाधारी) प्राण्यांप्रमाणे ब्रॅकिओपोडाही सागरातील अनुगमीर (खोलीनुसार असणाऱ्या प्राण्यांच्या वितरणाच्या बॅथिमेट्रिक) दृष्टीने महत्त्वाचे आहेत. [→ ब्रॅकिओपोडा].
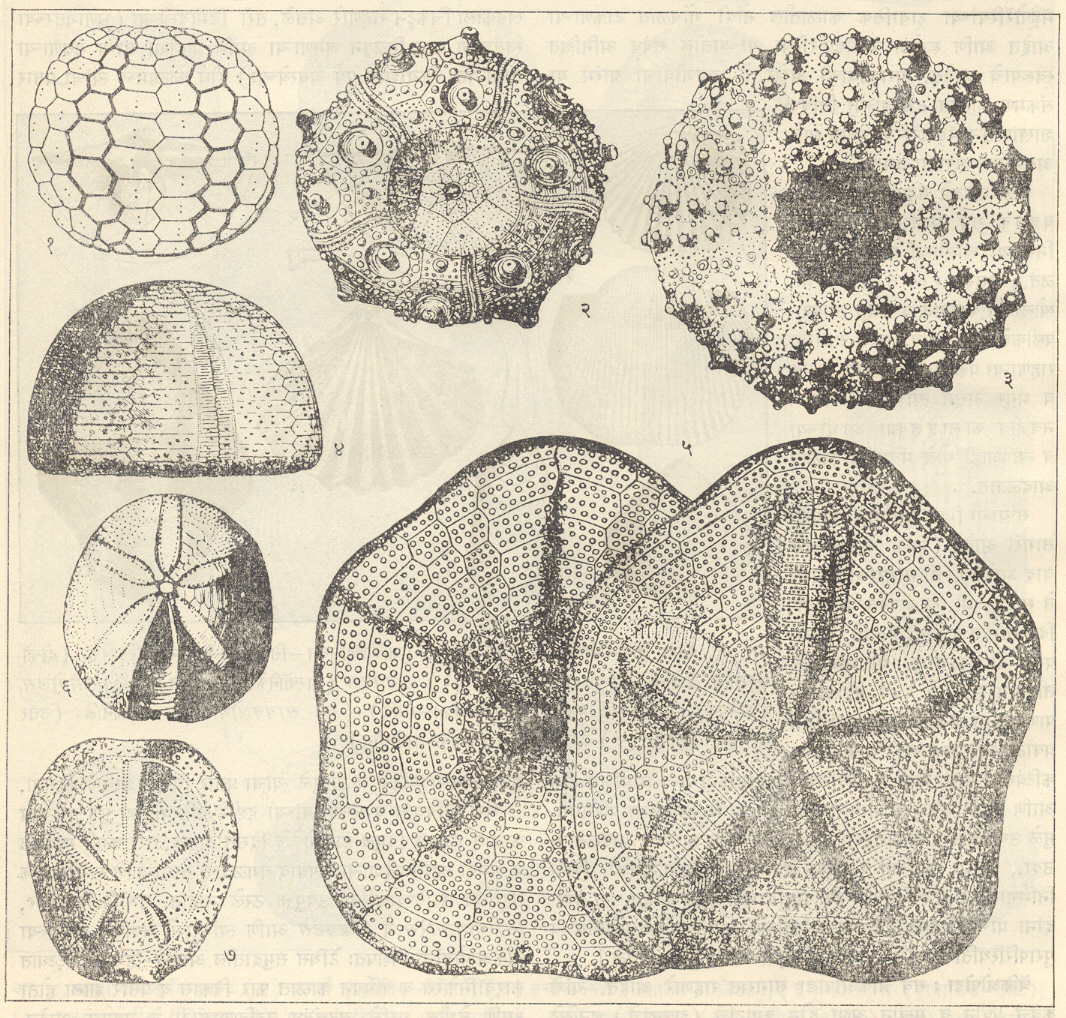
एकिनॉयडीया : (एकायनोडर्माटा). आधुनिक एकायनोडर्म (रेग्युलॅरिया) उदा., सिडारिस प्राण्याचे कवच गोलसर असून ते परिवलयाचे बनलेले असते. एकाआड एक अशी पाच चरणार-क्षेत्रे (ज्यांवर नालपाद-नळीसारखे पाय-असतात अशी क्षेत्रे) व पाच अंतराचरणार-क्षेत्रे यांच्यात मांडल्या गेलेल्या काहीशा षट्कोणी अशा अनेक तकटांचे परिवलय बनलेले असते. प्रत्येक क्षेत्रामध्ये तकटांचे दोन स्तंभ असून ते खालच्या पृष्ठाच्या मध्यावरील परिमुखापासून वरच्या शीर्षचक्रिकेपर्यंत पसरलेले असतात. शीर्ष चक्रिकेवर एकाआड एक पाच जननांग पट्ट व पाच अक्षीपट्ट (दृष्टीशी निगडित असलेले पट्ट) असून मध्यभागी परिगुद असते. अशा प्रकारे कवचाची सममिती पंचतयी (पाच सारख्या भागांनी तयार झालेली) असते. चर्वणाशी संबंधित असलेला ‘ॲरिस्टॉटलचा कंदील’ कवचाच्या आत परिमुखीय छिद्रालगत असतो व त्याच्याभोवती परिहु-मेखला (वलय अथवा कडे) असून तिला वरील कंदिलाचे स्नायू जोडलेले असतात. [→एकिनॉयडिया].
इंग्लंडमधील ऑर्डोव्हिसियन कालीन खडकांतील बोथिओसिडारिस (बोथ्रिओसिडारॉइडा गण) व ऑस्ट्रियातील उत्तर ट्रायसिक कालीन खडकांतील टायरेकिनस (प्लॅसिओसिडारॉइडा गण) हेच तेवढे असे माहीत असलेले प्राचीन एकिनॉयडीया प्राणी आहेत की, त्यांना आधुनिक एकिनॉयडियाप्रमाणे चरणार तकटांचे दोन स्तंभ आहेत परंतु बोथ्रिओसिडारिसमध्ये अंतराचरणार तकटांचा एक स्तंभ व टायरेकिनसमध्ये तीन स्तंभ होते. त्यांच्या परिहनु-मेखला व ॲरिस्टॉटलचा कंदील या अवयवांसंबंधी काहीच माहिती नाही.
प्राचीन एकिनॉयडीया गटातील दोन गणांपैकी एकायनोसिस्टाँयडा हा गण फक्त सिल्युरियनमध्येच आढळतो. यात चरणार तकटांचे २ ते ४ स्तंभ असतात परंतु अंतराचरणार तकटांची मांडणी उदग्रऐवजी क्षितिजसमांतर स्तंभांच्या रूपात झालेली असते. अशा तऱ्हेरने कवचाची सममिती पंचतयी नसते. पेरिशोएकिनॉयडा गणाच्या प्राण्यांमधील चरणार व अंतराचरणार तकटांची संख्या २ ते २० असते व कवचाची सममिती पंचतयी असते. हे प्राचीन एकिनॉयडियाचे प्रमुख गट असून हे सिल्युरियन ते पर्मियनअखेरपर्यंत होते आणि आर्किओसिडारिस, मीक्रेकिनस, हायटेकिनस इ. त्यांच्यातील विपुलपणे आढळणारे वंश होत. वरील दोन्ही गणांमध्ये बाजू कललेल्या असलेला ॲरिस्टॉटलचा कंदील असतो पण परिहनु-मेखला नसते.
आधुनिक एकिनॉयडियांपैकी सिडारॉइडा गणाची सुरुवात कार्बॉनिफेरस काळातील मायोसिडारिस वंशापासून झाली व सिडारिस हा त्याचा प्रातिनिधिक वंश होय. त्याच्या प्रत्येक अंतराचरणार तकटावर मोठी गुलिका असते व तिच्याभोवती कणिका असतात वा नसतात. शिवाय त्याला वर्तुळाकार परिमुख, अंतर्गत क्लोम (कल्ल्यासारखे अवयव) व मूळची परिवलयी तकटे वरच्या बाजूस वळून बनलेल्या परिहनु-मेखलेला क्रियाकारी स्नायूंनी जोडल्याने उदग्र झालेला ॲरिस्टॉटलचा कंदील ही असतात. काहीसा अपरिवर्तनशील असलेला हा गण आतापर्यंत हळूहळू बदलत गेला असल्याने त्यापासून फारसे परिवर्तित प्रकार निर्माण झालेले नाहीत.
ट्रायसिकच्या सुरुवातीस सिडारॉइडा गणापासून आलेला डायडेमॉइडा गण अतिशय जोमदार होता. त्यात परिहनु-मेखलेच्या अडथळ्यामुळे परिमुखीय क्लोम बाहेरील बाजूला गेले होते. परिणामी परिमुख दशकोनी झाले. दोन वा अधिक आद्य तकटे एकजीव होऊन चरणार तकटे संयुक्त होऊ लागली, अंतराचरणार तकटांची उंची कमी झाली आणि सर्व तकटांवर गुलिकांच्या कित्येक ओळी निर्माण झाल्या. परिवलयाची सममिती पंचतयी होती पण परिगुद पाठीमागे (कडेला) सरकल्याने शीर्ष चक्रिकेची सममिती द्विपार्श्व झाली. फायमोसोमा, डायप्लोपोडिया, स्यूडोडायाडेमा, सॅलेनिया, हेमिसिडारिस हे या गणातील काही प्रातिनिधिक वंश आहेत. हा गण विशेष करून जुरासिक व क्रिटेशस काळांत अतिविपुल होता. सिडारॉइडा व डायडेमॉइडा यांना मिळून रेग्युलॅरिया म्हणतात. त्यांचा नवजीव महाकल्पात काहीसा ऱ्हास झाला व ते हल्ली टिकून राहिलेले आहेत.
जुरासिकच्या प्रारंभी होलेक्टिपॉयडानंतर न्यूक्लिओलिटॉइडा व कॅसिडयुलॉइडा, पूर्व क्रिटेशसमध्ये स्पॅटँगॉयडा आणि उत्तर क्रिटेशसमध्ये क्लिपिॲस्ट्रॉयडा हे गण अवतरले. या सर्वांचा मिळून इर्रेग्युलॅरिया गट होतो. त्यांची वैशिष्ट्ये पुढील होत : लंबगोलाकार ते बदामासारखा आकार, कधीकधी पंचकोनी व द्विपार्श्व सममिती, खालचे पृष्ठ किंचित बहिर्गोल ते सपाट असलेला सामान्य शंक्वाकार वा अर्धगोलाकार, सर्व पृष्ठांवरील गुलिका असंख्य व लहान, शीर्ष चक्रिका जवळजवळ मध्याशी व परिमुख जवळजवळ मध्याशी किंवा काही स्पॅटँगॉयडामध्ये (उदा., हेमिॲस्टर, शिझॅस्टर, एपिॲस्टर इ.) अग्राकडे सरकलेले असते. चरणार क्षेत्रे होलेप्टिपॉयडामध्ये साधी, पण न्यूक्लिओलिटॉइडामध्ये पाकळीसारखी आणि स्पॅटँगॉयडामध्ये फक्त युग्मित चरणार क्षेत्रे पाकळीसारखी आणि स्पॅटँगॉयडामध्ये फक्त युग्मित चरणार क्षेत्रे पाकळीसारखी (उदा., हेमिॲस्टर, शिझॅस्टर), माक्रॅस्टरमध्ये काहीशी पाकळीसारखी (उदा., मायक्रॅस्टर) किंवा साधीही (उदा., होलॅस्टर) आढळतात, मात्र अयुग्मित अग्र चरणार क्षेत्र या सर्वांमध्ये साधे असते. वरच्या पृष्ठावरील चरणार स्पर्शकाद्वारे हे श्वसन करतात. त्यांच्यात परिमुखिय क्लोम, ॲरिस्टॉटलचा कंदील व परिहनु-मेखला नाहीशी झालेली असतात फक्त क्लिपिॲस्ट्रॉयडा व काही होलेक्टीपॉयडा यांमध्येच ऱ्हासपावलेली मेखला व कंदील टिकून राहिलेली असतात. शीर्ष चक्रिका व परिरेखा (शीर्षाकडून पाहिले असता दिसणारी कवचाची बाह्य रेखा) यांच्या दरम्यान परिगुद असते व कोन्युलससारख्या काहींच्या बाबतीत ते परिरेखेच्याही खाली असते.
हे सर्व गण जुरासिक व क्रिकेटशसमध्ये विपुल होते आणि आता समुद्रात राहत आहेत. स्पॅटँगॉयडा गण नवजीव महाकल्पात विपुल होता. दीर्घकाल इतिहास असलेले एकिनॉयडिया सामान्यपणे थरांचे सहसंबंध स्थूलपणे ठरविण्यास उपयुक्त आहेत परंतु मायक्रॅस्टरसारखे काही एकिनॉयडीया जीवाश्म कालदर्शक आहेत, तर काही बाबतींत मर्यादित क्षेत्रांतील शैलसमूहांचे उपविभाग पाडण्यास ते उपयुक्त ठरु शकतात.
एकिनॉयडिया बहुतकरून उष्ण आणि उपोष्ण कटिबंधीय असून समुद्रतटीय व उपसमुद्रतटीय पट्ट्यात ते विपुल असतात, त्यांच्या काही जाती अधिक खोल पाण्यातही आढळतात. एकिनॉयडिया हे जात्याच नितलस्थ प्राणी असून ते सामान्यपणे गाळवटी व चिखलयुक्त पाणी टाळतात मात्र क्लिपिॲस्टर, एंकोप, एकायनोकार्डियम, स्पॅटँगस इत्यादींच्या काही जाती खडकांत बिळे करूनही राहतात. [→ एकिनॉयडीया].
बायव्हाल्व्हिया : (मॉलस्का). याला लॅमेलिब्रँकिया, पेलिसिपोडा किंवा शिंपाघारी वर्ग असेही म्हणतात. ट्रायलोबिटा, ब्रॅकिओपोडा, स्पंज इ. केवळ सागरी प्राण्यांच्या अवशेषांबरोबरच बायव्हाल्व्हियांचेही सुरुवातीचे अवशेष सागरी खडकांत आडकांत आढळतात. त्यावरून बायव्हाल्व्हियांचीही उत्पत्ती सागरी पर्यारवरणात झाली, हे उघड आहे. ते आतापर्यंत मुख्यत्वे सागरी प्राणी आहेत पण गॅस्ट्रोपोडाप्रमाणे त्यांनी मचूळ व गोड्या पाण्याच्या परिस्थितीशीही जुळवून घेतले आहे. अँथ्रॅकोमाया आणि कार्बॉनिकोला हे कार्बॉनिफेरस काळातील वंश गोड्या पाण्यातील सर्वात आधीच्या बायव्हाल्व्हियांच्या वंशापैकी आहेत.
तळावरील गाळातून नांगराप्रमाणे उरकत जाऊन आपला मार्ग काढणारे बायव्हाल्व्हिया हे मुख्यत्वे नितलस्थ प्राणी असून त्यांतील काही समुद्रतळानजीक पोहू लागले आणि उघड्या सागरांत काही तलप्लावी (मध्यम खोलीवर व पृष्ठालगतच्या पाण्यात राहणारे) व प्लवकजीवरूपीही (पाण्यावर तरंगणाऱ्या प्राण्यांच्या प्रकारचेही) झालेले आढळतात. पुष्कळ नितलस्थ गटांनी आपली जीवनप्रणाली बदलली व ते तात्पुरते वा कायमचे स्थायी (स्थिर) प्राणी बनले काही अधिक खोल जागी जाऊन जमीन उकरून मार्ग काढण्याची जीवनप्रणाली अधूनमधून अनुसरतात. फोलॅडोमायासारख्या काही
प्रकारांत शिंपा पूर्णपणे न मिटता त्यांच्या पश्च भागी फट निर्माण झाली. काही तळावरील खडकांत बिळे करून राहू लागले. उदा., लिथोफॅगस चुनकडकांत तर ⇨ फोलॅस व सॅक्सिकाव्हा टणक वाळूत आणि ⇨ टेरेडो पाण्यातील झाडांच्या खोडांत वा गोदीमधील बांधकामातील लागडांत छिद्रे पाडतात. ⇨ ऑयस्टर आपले कवच (शिंपा) सरळसरळ कठीण खडकावर चिकटवितात व त्यातून ऑयस्टर किनारे तयार होतात. पिनिडी, मायटिलिडी, मॉडिओलिडी, टेरिइडी या कुलांतील प्राणी सूत्रगुच्छ रज्जूने (चिवट तंतूंच्या जुडग्याने) तळावरच्या खडकाला जखडले असतात.

पेक्टिनिडीमधील प्राणी प्रथम असेच जखडवून घेणारे होते पण नंतर ते मुक्त पोहणारे झाले. मूलतः बायव्हाल्व्हियांच्या शिंपेला पश्च स्नायूपेक्षा किंचित लहान अग्रीय स्नायू असलेला अग्र भाग होता. विविध प्रकारच्या जीवनप्रणालींमुळे शिंपेत परिवर्तने होतात. अशा प्रकारे तळावरील गाळातून उकरत जाण्यामुळे शिंपेच्या अग्र भागाची तसेच त्याच्या जोडीने अग्रीय स्नायूची वाढ मर्यादित होते. त्यामुळे असममिती वाढत जाऊन काही जातींच्या बिळे करण्याच्या प्रवृत्तीमुळे ती अतीशय वाढते व फोलॅसप्रमाणे शिंपा जवळजवळ दंडगोलाकार होते. जखडून राहणाऱ्या प्रकारांत अग्र बाजूवर सारखी ओढ पडत असल्याने शिंपेची अग्र बाजू व तिच्या बरोबर अग्रीय स्नायूचा ठसा वाढत्या प्रमाणात अस्पष्ट होत जाऊन शेवटी शिंपा पाचरीसारखी होते व अग्रीय स्नायूच्या ठशाचा ऱ्हास होत जातो उदा., मायटिलस, टेरीया, पिन्ना वगैरे. ऑयस्टरांमध्ये सामान्यपणे डावी शिंपा सरळ तळाच्या खडकांवर तयार होते व तिचा बाह्याकार अतिशय ओबडधोबड होतो. सुट्या शिंपेचा मूळचा बाह्याकार पुष्कळसा टिकून राहिलेला असून ती केवळ झाकण्यासारखी असते. फक्त पश्च स्नायू प्रत्यक्ष कार्यक्रम असतो व बिजागरीचे काहीच काम उरत नसल्याने ती पूर्णतया ऱ्हास पावते. शिंपेमधील या बदलांबरोबर बिजागरीचा अग्र भाग दबला जाऊन विरल-दंती व इतर कित्येक वैविध्यपूर्ण विषम-दंती बिजागरी बनते. त्याचप्रमाणे ज्यांचा अग्र स्नायू अधिकाधिक दुर्बल झालेला आहे, अशा द्वि-अभिवर्तनी (दोन स्नायु-ठसे असलेल्या) शिंपांपासून एक-अभिवर्तनी (एक स्नायु-ठसा असलेल्या) शिंपा निर्माण होतात. ग्रिफीया व एक्झोगायरा आणि त्यांच्याशी निगडित असलेल्या प्राण्यांमध्ये वरची शिंपा सपाट किंवा अंतर्गोलही असते, मात्र खालची शिंपा अतिशय बहिर्गोल झालेली असून ककुद (चोचीसारखा दिसणारा भाग) पिळवटलेला वा कुंडलित असतो. रूडिस्टेसमध्ये न हलणारी (अचल) शिंपा उलट्या शंकूप्रमाणे वरच्या दिशेत वाढते व ती पोवळ्यासारखी दिसते आणि मुक्त शिंपा ही केवळ झाकण म्हणून असते व तिचा मूळ आकार जसाच्या तसा टिकून राहिलेला असतो. हिप्प्युराइट व रेडिओलाइट ही यांची नमुनेदार उदाहरणे होत.
या विविध जीवनप्रणाली सभोवतालच्या परिस्थितीशी निगडित असतात आणि त्या पर्यावरणात राहणाऱ्या शिंपांची सर्वसाधारणतः आकार व इतर वैशिष्ट्ये महत्त्वाची असतात. अशा प्रकारे जे बायव्हाल्व्ह प्राणी तळावरील गाळावर रांगतात किंवा तो उरकत पुढे जातात त्यांच्या दोन्ही शिंपा एकसारख्या बहिर्गोल असून त्यांचा कमाल बहिर्गोलपणा सामान्यपणे उंचीच्या मध्याच्यावर असतो आणि गाळ कापीत पुढे जाणे सुलभ व्हावे म्हणून शिंपांच्या कडा पाचरीच्या आकाराच्या झालेल्या असतात. पेक्टेन, इनोसेरॅमस इत्यादींप्रमाणे जे बायव्हाल्व्ह एका वा दुसऱ्या बाजूवर तात्पुरते विसावतात, त्यांची एक शिंपा दुसरीपेक्षा अधिक बहीर्गोल असते. जोरदार प्रवाह असणाऱ्या भागात राहणारे बायव्हाल्व्ह सूत्रगुच्छाद्वारे तळाला जखडून राहतात, खडकाळ किंवा अन्यथा कठीण तळ असणाऱ्या भागातील बायव्हाल्व्ह ऑयस्टराप्रमाणे आपल्या शिंपा दृढ खडकाला संयोजक द्रव्याने सरळ चिकटवून टाकतात (ऑयस्टर समुद्रतटीय व उपतटीय पट्ट्यात असेच चिकटून राहिलेले असतात व तेथे शिंपायुक्त विस्तृत किनारे निर्माण झालेले असतात). पेक्टिनिडी व ग्लायसेमेरिस यांच्यासारख्यांच्या जवळजवळ वर्तुळाकार शिंपा मुक्तपणे पोहण्यास सोयीच्या असतात. पातळ, सपाटसर व जवळजवळ सममित असलेल्या शिंपा आणि तरंगण्याची जीवनप्रणाली एकमेकांशी संबंधित असतात. निनाल (नलिकाकार अवयव). सभोवतालच्या पाण्याबाहेर ठेवून प्राणी तळावरील गाळात बीळ करीत गेल्याचे पश्च फट असलेल्या शिंपावरून दिसून येत. समुद्रतटीय भागात जे बायव्हाल्व्हिया तळाला न चिकटता राहतात त्यांची शिंपा गोल दंडाकार, जड असते कारण या जडपणामुळे येथील क्षुब्ध स्थितीत लाटेबरोबर लोळण्यास सुलभ असल्याने ती तग धरून राहते खंड-फळीवरील (खंडाच्या किनाऱ्यालगतच्या कमी उताराच्या भागावरील) अधिक खोल भागात व त्यापलीकडील भागात अन्न मिळविण्याच्या स्पर्धेत शिंप्यांच्या जडपणाचा अडथळा नको म्हणून त्या पातळ व हलक्या बनलेल्या असतात.
बायव्हाल्व्हिया कँब्रियन कल्पाच्या आधीच्या काळात अवतरले व नंतर ऑर्डोव्हिसियन व सिल्युरियनपर्यंत त्याचा जलद विकास झाला परंतु उत्तर पुराजीव काळात त्यांची वाढ खास डोळ्यांत भरण्याइतकी झाली नाही.
पुराजीवकालीन बिजागरी असलेले प्रकार हे दात व खोबणी स्पष्ट नसलेले आदिम पॉलिओकाँच (पुरावंश) होते. त्यांच्या टेनोडोंट, ॲक्टिनोडोंट व टॅक्सोडोंट (बहुदंती) या सर्व वंशजांमध्ये अविभेदित अनेक दात व खोबणी होत्या. प्रमुख बदल बहुतकरून ॲक्टिनोडोंटमध्ये झाला व तो म्हणजे दातांची संख्या घटणे व ते बळकट होणे हा होय. युनिऑड बिजागरी डेव्होनियन काळात प्रस्थापित झाली. तिच्यात युनिओ, कार्डिनिया, अँथ्रॅकोमाया, कार्बॉनिकोला इत्यादींप्रमाणे ४ ते ५ दात असतात. ककुदाखालील आणखी दोन दातांची घट झाल्याने दोन संभाव्य दिशांत विकास झाला. अप्रगतशील दिशेने झालेल्या विकासात ट्रयगोनियाइडीप्रमाणे विरल-दंती बिजागरी आढळते. दुसऱ्या दिशेतील विकासामध्ये मूळ दातांच्या दोन्ही बाजूंना ककुदाखाली एक किंवा दोन पार्श्वीय दातांची (यांना प्रमुख दात म्हणतात) भर पडली आणि त्यापासून ट्रायासिकमध्ये प्रगतशील विषमदंती (किंवा विविध-दंती) बिजागरी असलेले प्रकार निर्माण झाले व त्यांच्यापासून मध्यजीव व नवजीव काळांत कित्येक विविधतापूर्ण प्रकार निर्माण झाले. ही बिजागरी असलेला गट हल्ली प्रमुख आहे. अर्थात टेक्सोडोंट, अक्सिनोडोंट व टेनोडोंट हे अविशेषित प्रकारही हल्ली अस्तित्वात आहेत.
मेगॅलोडोंटिडी, ट्रायगोनिइडी, पिनिडी, पेक्टिनिडी, मायटिलिडी इ. बायव्हाल्व्हियांची मचूळ पाण्यातील कुले डेव्होनियनमध्ये अवतरली व तेव्हापासून आजपर्यंत अखंडपणे ती विपुल प्रमाणात टिकून राहीलेली आहेत. ट्रायगोनिइडी हे कुल याला अपवाद असून ते जलदपणे ऱ्हासपावत आहे. यापेक्षाही दीर्घ काळ असलेली दोन कुले म्हणजे ऑर्डोव्हिसियनपासून आतापर्यंत असलेले लेडिडी व सिल्युरियनपासून आतापर्यंत असलेले न्यूकुलिडी ही होत परंतु या दीर्घ काळात त्यांच्यात फारच थोडा बदल झालेला दिसून येतो. ट्रायासिस अखेरच्या खंडानंतर जुरासिक व पूर्व क्रिटेशसमध्ये बायव्हाल्व्हिया अतिशय विपुल व विविध प्रकारचे होते. तेव्हापासून गॅस्ट्रोपोडांची संख्या त्यांच्यापेक्षा जास्त झाली. मात्र असे असले, तरी हल्लीच्या मॉलस्का जीवांमधील बायव्हाल्व्हिया हा महत्त्वाचा घटक आहे.
बायव्हाल्व्हियांची बहुतेक कुले दीर्घकालीन आयुर्मर्यामदेची असल्याने स्तरवैज्ञानिक सहसंबंध निश्चित करण्याच्या दृष्टीने त्यांची उपयुक्तता कमी आहे. तथापि पुष्कळ वंशांची अशी चांगलीच उपयुक्तता आहे उदा., पर्मियन व कार्बॉनिफेरसमध्ये मेगॅलोडोंट पर्मीयनमध्ये ॲव्हिक्युलोपेक्टेन विशेषतः जुरासिक व क्रिटेशसमध्ये ट्रायगोनिया जुरासिकमध्ये ग्रिफीया उत्तर जुरासिक व क्रिटेशसमध्ये एक्झोगायरा पूर्व क्रिटेशस, ऑलिगोसीन (सु. ३.५ ते २ कोटी वर्षांपूर्वीचा काळ) व मायोसीन (सु. २ ते १.२ कोटी वर्षांपूर्वीचा काळ) यांमध्ये ऑस्ट्रियाच्या काही जाती आणि क्रिटेशसमध्ये रेडिओलाइट, हिप्प्युराइट, इनोसोरऐमस, नायथिमा, मायटिलस व पिन्ना.
जीवाश्मरूप शिंपांमधील बिजागरीची वैशिष्ट्ये, स्नायु-ठसे व प्रवार-रेखा यांच्या आधारे एम्. न्यूमायर व वुड्स यांनी वर्गीकरणाची पद्धती तयार केली. या पद्धतीने क्रमसूचक पातळीवर पडणारे विभाग जिवंत बायव्हाल्व्हयांच्या मऊ भागांना अनुसरून केलेल्या प्राणिवैज्ञानिक वर्गीकरणाशी बहुतकरून जुळतात. [→बायव्हाल्व्हिया].
गॅस्ट्रोपोडा : (मॉलस्का). गॅस्ट्रोपोडाचे कवच म्हणजे शंख होय व तो मूलतः शंक्वाकार असतो. त्याची सुरुवात पूर्व कँब्रियन काळातील स्टेनोथिका, स्केनेला, पॅलॅक्मिया इत्यादींमधील बसकट टोपीसारख्या शंखाने झाली. या टप्यानंतर मध्य पुराजीव काळातील बेलेरोफोन व त्याच्याशी संबंधित गॅस्ट्रोपोडांच्या सपाट-सर्पिल कुंडलाकार शंखाचा टप्पा आला. मळसूत्रज कुंडल हा यानंतर झालेला संरचनात्मक विकास असून मळसूत्रज कुंडल हे शंखाचे सर्वांत सामान्यपणे आढळणारे स्वरूप आहे. शंखांचा आकार सामान्यपणे बिंबाभापासून (उदा., प्लॅनॉर्बिस) ते अतिशय निमूळता घुमटाकार (उदा., टरिटेला, एरिथियम, नेरिनिया) अशा व्यापक आकारमालिकेत आढळतो. शंखाचा आकार पुढीलप्रमाणेही असू शकतो : नॅटिका व टर्बोमध्ये गोलाकार, ट्रॉकस व प्लुरोटोमॅरियामध्ये शंक्वाकार, कोनसमध्ये द्विशंक्वाकार, फ्यूसस आणि म्यूरेक्समध्ये चातीच्या आकाराचा, सायप्रियामध्ये संवलित, मर्चिसोनिया व लोक्सोनिमामध्ये बुटक्या मनोर्यामप्रमाणे. शंख बहुतकरून दक्षिणावर्ती (उजव्या तोंडाचा) कुंडलित असून फायसा व प्लॅनॉर्बिसच्या काही जातींमधल्याप्रमाणे काही वामावर्ती (डाव्या तोंडाचा) प्रकारही माहीत आहे पण ते शंख विरळाच आढळतात. शंखामध्ये विकुंडलनाची (वलयांचा ऱ्हास होण्याची) प्रवृत्तीही आढळते. उदा., स्कॅफँडर, क्रेपिड्युला, हेलिओटिस, कॅलिप्ट्रिया, व्हर्मिट्स इत्यादी. वरवर पाहता पटेलांचा शंख जरी एखाद्या साध्या आद्य शंकूसारखा दिसत असला, तरी तो पूर्णपणे विकुंडलन झालेला शंख आहे.
स्पंज, ट्रायलोबाइट, ग्रॅप्टोलाइट, ब्रॅकिओपोडा इ. सर्वस्वी सागरी प्राण्यांच्या जोडीन आढळणारे गॅसट्रोपोडा हे सागरी जीव म्हणून कँब्रियनच्या प्रारंभी अवतरले. मुख्यत्वे सागरी असलेल्या गॅस्ट्रोपोडांनी मचूळ व गोडे पाणी या अधिवासांशी जुळवून घेतले आहे, तसेच विविध जलवायुमानीय (दीर्घकालीन सरासरी हवामानाची) स्थिती असलेल्या जमीन या अधिवासाशीही त्यांचे अनुकूलन झाले आहे. डेव्होनियन काळातील डेंड्रोप्युपा हा गोड्या पाण्यातील सर्वांत आधीच्या गॅस्ट्रोपोडांपैकी एक वंश असून फायसा व अँप्यूलॅरिया यांचे आधीचे नातेवाईक हे कार्बॉनिफेरस काळातील गोड्या पाण्यातील गॅस्ट्रोपोडा होत.
सागरी पर्यावरणामध्ये गॅस्ट्रोपोडा हे मुख्यत्वे सागरी तळावर रांगणारे व तळावरील गाळ उरकत पुढे जाणारे नितलस्थ प्राणी आहेत. सागराच्या तळाजवळ ते पोहतात व उघड्या समुद्रात ते तलप्लावी व प्लवकजीव म्हणूनही आढळतात. जीवनप्रणालीला अनुरूप असा जड किंवा हलका शंख निर्मिण्याची त्यांची प्रवृत्ती असते, तर हल्लीच्या न्यूडिब्रँकियांना शंखच नसतो.
सागरी पर्याणवरणामध्ये गॅस्ट्रोपोडा समुद्रतटीय पट्ट्यात अतिशय विपुल असून भरती-ओहटीच्या व प्रवाहांच्या तेथील जोरदार हालचालींमध्ये टिकून राहण्याच्या दृष्टीने त्यांचे शंख जड व बळकट झालेले असतात. नॅसा, नॅसारिया, लिटोरिना, नेपच्यूनिया, स्ट्रॉँबस हे येथील सामान्य प्रकार असून खडकाला चिकटून राहणाऱ्या लिंपेट गटाच्या पटेलासारख्या प्रकारांचा शंख टोपीसारखा असतो व त्यामुळे लाटांनी ते वाहून नेले जात नाही. खंड-फळीवरील वरच्या सु.१५ ते ५० मी. खोलीपर्यंतच्या पट्ट्याला लॅमिनारियन व सु. ५० ते १०० मी. खोलीपर्यंतच्या पट्ट्याला नलिपोर पट्टा म्हणतात. तेथे जीवांची अतिशय गर्दी असून त्यांच्यात जीवन स्पर्धा फार तीव्र असते. अशा ठिकाणी अन्न मिळविण्याच्या स्पर्धेत जड शंख हा भारच होईल, तसेच या भागात लाटा व प्रवाह यांचे परिणाम फारच कमी होतात. त्यामुळे येथील गॅस्ट्रोपोडांचे शंख हलके बनलेले असतात. येथील सामान्य वंश पुढील होत : नॅसा, ट्रॉकस, रिसोला, टरिटेला, नॅटीका, बक्सिनम इत्यादी. येथे वारंवार आढळणाऱ्या प्राण्यांच्या प्रकारांमध्येही वाळू व चिखल यांविषयीची आवडनिवड स्पष्टपणे दिसू येते. खंड-फळीच्या पलीकडच्या वितलीय (साधारण २,००० ते ६,००० मी. खोलीच्या) भागात जीवांचे प्रमाण जलदपणे घटत जाते व येथील गॅस्ट्रोपोडांचे शंख अधिक पातळ व रंगहीन असतात.

गॅस्ट्रोपोडा जरी अतिशय विविध प्रकारच्या संजातीचे असले, तरी त्यांचे शंख ज्या परिस्थितीत ते राहतात त्या परिस्थितीला अनूरूप अशा सामान्य आराखड्याचे असतात. म्हणून या दृष्टीने अभ्यास केला, तर शंखांना पर्यावरणीय महत्त्व आहे.
गॅस्ट्रोपोडांची संख्या व विविधता पुराजीव कल्पभर सावकाश पण सतत वाढत होती तरी प्रावारात पाणी येण्याजाण्याच्या मार्गासाठी असलेली प्रावारातील फट आणि तिच्याशी संबंधित अशी शंखांमधील द्वारकाच्या बाह्य ओष्ठातील खोबण (उदा., बेलेरोफोन, लोक्सोनिमा, प्ल्युरोटोमारिया, मर्चिसोनिया, यूओफॅलस इत्यादींमधील खोबणी) निर्माण होण्याच्या टप्प्यापलीकडे त्यांची प्रगती झालेली नव्हती. सध्याच्या सागरी कुलांपैकी ट्रोकिडी, लिटोरिनिडी, टर्बिनिडी, एपिटोनिइडी, टरिटेलिडी ही कुले उत्तर पुराजीव काळात अवतरली आणि ट्रायासिक काळानंतर त्यांची भरभराट झाली. ज्यांच्यात अग्र व पश्च नाल चांगले विकसित झाले आहेत असे सायफोनेट (निनालयुक्त) गॅस्ट्रोपोडा ट्रायासिकमध्ये आढळतात व तेव्हापासून गॅस्ट्रोपोडांतील हा प्रमुख गट बनलेला आहे.

सामान्यपणे दीर्घकालीन आयुर्मर्यादा असलेले गॅस्ट्रोपोडा स्तरवैज्ञानिक सहसंबंधांमध्ये उपयुक्त नसतात. तथापी काही वंश स्थानिक वा प्रादेशिक दृष्ट्या सूचक जीवाश्म म्हणून उपयुक्त असल्याचे आढळले आहेत. उदा., लोक्सोनिमा, प्ल्युरोटोमारिया, मर्चिसोनिया, यूओंफॅलस हे उत्तर पुराजीव काळात, अँथलिटा उत्तर जुरासिक काळात, फायसा उत्तर क्रिटेशस व इओसीन (सु. ५.५ ते ३.५ कोटी वर्षांपूर्वीच्या) काळात, गायसॉर्टीया इओसीनमध्ये, नेरिनिया उत्तर क्रटेशसमध्ये, गॅस्ट्रोपोडा बायव्हाल्व्हियांबरोबर अवतरले आणि त्यांच्याप्रमाणे गॅस्ट्रोपोडांची संख्या व विविधता यांत वाढ झाली परंतु मध्य क्रिटेशसरर्यंत गॅस्ट्रोपोडा बायव्हाल्व्हियांच्या मानाने गौण होते. तथापि तदनंतर गॅस्ट्रोपोडा मॉलस्कातील प्रमुख प्राणी होत गेले. नवजीव हा त्यांच्या विकासातील परमोत्कर्षाचा काळ असून प्लायोसीन (सु.१.२ कोटी ते ६ लाख वर्षांपूर्वीच्या) काळातील यांच्या ९० टक्क्यांपेक्षा जास्त जाती असून अजूनही अस्तित्वात आहेत.
गॅस्ट्रोपोडा मुख्यत्वे उष्ण पाण्यात राहणारे असून जमिनीवरील व महासागरातील विशेषतः सध्याचे प्राणिजातींचे प्रदेश निश्चित करण्यासाठी त्यांची मदत होते. जुरासिक, क्रिटेशस व नवजीवन काळांतील गॅस्ट्रोपोडांची वाटणी पाहिल्यास ती अधिकाधिक प्रमाणात प्लायोसीन अखेरीस प्रस्थापित झाली होती. ती आजच्या त्यांच्या प्रदेशासारखी होत गेलेली दिसते.
तंत्रिका वलयाचे (मज्जा-वलयाचे) स्वरूप, हृदयाचे स्थान व क्लोमांचे (कल्ल्यांचे) स्वरूप यांच्यावर गॅस्ट्रोपोडांचे मुख्य वर्गीकरण आधारलेले असते. त्यांपैकी कोणाचाच शंखांशी प्रत्यक्ष संबंध येत नाही. रेत्रिका (कायटिनी दातांच्या ओळी असलेली पट्टी) व प्रावारातील फटीचा विकास तसेच शंखांच्या मुखाची बाह्यरेखा बदलणाऱ्या पाण्याच्या मार्गासाठी असलेला निनाल आणि शंख न बनविण्याची प्रवृत्ती हे घटक गौण पातळीवरील मानले जातात. म्हणून गॅस्ट्रोपोडांच्या जीवाश्मांचा विचार करताना शंखाची सर्वसाधारण वैशिष्ट्ये आणि त्यावरील नक्षीकाम यांचा उपयोग वंश व जाती व या पातळ्यांवर काही अंशी कुलाच्या पातळीवर होऊ शकतो. [→ गॅस्ट्रोपोडा]
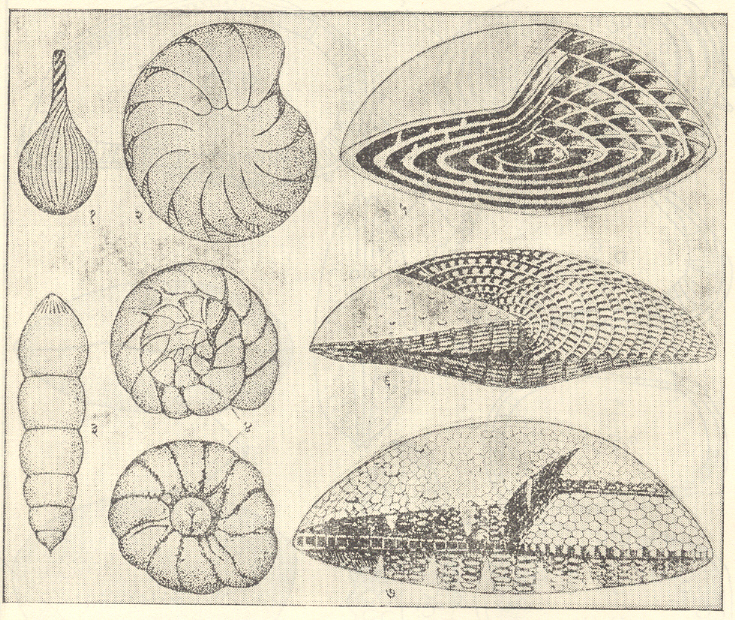
नॉटिलॉइडीया : (सेफॅलोपोडा मॉलस्का). नॉटिलॉइडिया गटाचे प्राणी ऑर्डोव्हिसियन कल्पाच्या प्रारंभी किंचित वक्र व आखूड शंकूच्या रूपात अवतरले. या गटातील ⇨ नॉटिलसच्या फक्त दोन (कदाचित तीन) जाती हल्ली हिंदी-पॅसिफिक महासागरांच्या उष्ण कटिबंधीय भागात अस्तित्वात आहेत व त्यामुळे त्यांचे वर्गीकरण करणे अवघड झाले आहे. तथापि त्यांचे सामान्यतः पुढील तीन उपवर्ग मानले जातात : (१) एंडोसेरॅटॉयडिया व (२) ॲक्टिनोसेरॅटॉयडिया : हे दोन्ही ऑर्थोकोनिक ते सिर्टोकोनिक असून क्वचित त्यांचे कवच रुंद निनालिका व अंतर्गत चूर्णीय निक्षेप आणि निनालिकीय संरचना असणारे सैलसर कुंडलित असे असते आणि (३) नॉटिलॉइडिया : या उपवर्गातील कवच ऑर्थोकोनिक ते सिर्टोकोनिक असून ते सैलसर वा घट्टपणे कुंडलित असते. निनालिका जलदपणे अरुंद होत गेलेली असते आणि मुख्यत्वे मध्य पुराजीव काळातील गटांत अंतर्गत निक्षेप व निनालिकीय संरचना आढळतात. शेवटचा उपवर्ग नॉटिलॉइडियाचा प्रमुख गट असून त्यांपैकी काहींमध्ये शंखाच्या पृष्ठभागावर ओबडधोबड घड्या, फासळ्या व गाठी या रूपांत नक्षी निर्माण झालेली असते. साधी सीता हे एकूण नॉटिलॉइडियांचे वैशिष्ट्य असून ती सरळ अथवा ऑर्थोकोनिक व सिर्टोकोनिक कवचांतल्याप्रमाणे किंचित वक्र असते व नॉटिलससारख्या कुंडलित प्रकारांमध्ये स्थूलपणे एकसारखे एक-दोनच साधे खंड व पल्याण (खोगिराच्या आकाराचे भाग) पडलेले असतात. ॲट्युरिया, ॲट्युरॉइडिया व हेर्कोग्लॉस या वंशांत खंड व पल्याण थोडेफार निमूळते असतात.
अरुंद कोटर, अंतर्गत व निनालिकीय संरचना यांच्यामुळे नॉटिलॉडियांच्या कवचाची उत्प्लावकतां (उद्धरणक्षमता) अल्प असते त्यामुळे ते मुख्यत्वे तरणक-नितवस्थ (सागराच्या तळालगत पोहणारे) असले पाहिजेत आणि जीवश्मांवरून दिसणारा त्यांचा विसतृत प्रसार हा बहुधा त्यांच्या मृत्यूनंतर झाला असला पाहिजे. त्यांच्यापैकी पुष्कळ प्राणी वाहून दूर नेले जाऊन त्यांचा मूळ अधिवास जेथे नव्हता अशा तेव्हाच्या एखाद्या द्रोणीमध्ये साचले असावेत. म्हणून ते स्तरांचे सहसंबंध केवळ स्थूलपणे ठरविण्यासाठीच उपयुक्त असू शकतील व त्यांच्यापैकी काही प्राणी मर्यादित क्षेत्रांतील सहसंबंधासाठी अधिक उपयोगी ठरू शकतील.
अधिक रुंद कोटर आणि अंतर्गत निक्षेप व निनालिकीय संरचना यांचा अभाव यांच्यामुळे अधिक लांब ऑर्थोसेरॅटिड व उत्तर पुराजीव काळातील पुष्कळ कुंडलित नॉटिलॉइडिया यांची कवचे उत्प्लावक होती व त्यामुळे हे प्राणी पोहणारे असले पाहिजेत म्हणून त्यांच्या विस्तृत प्रसारामुळे ते दूरदूरच्या विविध द्रोणींमधील थरांचे सहसंबंध ठरविण्यासाठी उपयुक्त ठरले आहेत.
मध्य पुराजीव काळातील फ्रॅग्मोसेरस, गाँफोसेरस इत्यादींसारख्या काही आखूड ऑर्थोकोनिक नॉटिलॉइडांमध्ये अनुकूलता परिणामी यांचे द्वारक (मुख) लहान झाले आहे व ते तरंगत राहणारे प्राणी असले पाहिजेत. लिट्युआइट आणि ट्रोकोसेरससारख्या काही नॉटिलॉइडांमध्ये विकुंडलनाची प्रवृत्ती दिसून येते. शेवटच्या मलसूत्रज कुंडलित प्रकारांपैकी काही बहुधा नितलस्थ होते.
बहुतेक नॉटिलॉइडिया डेव्होनियन कल्पाच्या शेवटीच नष्ट झाले, काहींची कार्बॉनिफेरस व पर्मियन काळांत भरभराट झाली. उदा., ऑर्थोसेरस, फूर्डिसेरस, मेटॅकोसेरस, टेम्नोकायलस, व्हेस्टिनॉटिलस. त्यांच्यापैकी बहुतेक पुराजीव काळाच्या शेवटी निर्वंश झाले. ऑर्थोसेरॅटिडींपैकी ऑर्थोसेरस व मिचेलिनोसेरस आणि टेनोसेरॅटिडी, ग्रिपोसेरॅटिडी व लायरोसेरॅटिडी हे ट्रायासिकमध्ये आलेले कुंडलित नॉटिलॉइडिया होत. ग्रिपोसेरस, टेनोसेरस, पॅरानॉटिलस, क्लायडोनॉटिलस, प्ल्युरोनॉटिलस व नॉटिलस या वंशांचा ट्रायासिक काळात विस्तृत प्रसार झालेला आहे. ट्रायासिकमध्ये अवतरलेला नॉटिलस वंश जुरासिक व क्रिटेशस काळांत जवळजवळ जगभर पसरला होता आणि पॅरासोनोसेरस, पॅरासायमॅटोसेरस, हेर्कोग्लॉसा, ॲट्युरॉइडिया व हेमिनॉटिलस हे क्रिटेशस काळांतील काही प्रमुख प्रकार आहेत. नवजीव महाकल्पाच्या आधिच्या काळात नॉटिलॉइडियांचा अतिशय जलदपणे ऱ्हास झाला. ॲट्युरिया, सायमॅटोनॉटिलस, हेर्कोग्लॉसा, डेल्टॉइडोनॉटिलस व नॉटिलस हे त्या काळातील सामान्यतः आढळणारे वंश होते.
चतुर्थ (गेल्या सु. ६ लाख वर्षांच्या ) काळात केवळ नॉटिलस वंश जिवंत राहीला . त्याच्या दोन (कदाचित तीन) जाती हल्ली हिंदी-पॅसिफिक महासागरांच्या उष्ण कटिबंधीय भागातील ईस्ट इंडीज ते फिजी बेटांपर्यंतच्य प्रदेशात आढळतात. [→ नॉटिलॉइडिया].
ॲमोनॉइडिया : (सेफॅलोपोडा मॉलस्का). सरळ कवच अललेले बॅक्ट्रायटिस किंवा सैलसरपणे कुंडलित असे बॅरँडीओसेरस ज्या नॉटिलॉइड शाखेतील होत तिच्यापासून सिल्युरियनअखेरीस किंवा डेव्होनियन काळाच्या सुरुवातीस ॲमोनॉइड अवतरले. त्यांचे प्रारंभीचे कवच साधी सीता असलेले, सैलसर कुंडलित, साधे व नितळ होते व सीतांचे स्थूलपणे सारखे दोन खंड (मागील दिशेत बाक असलेले) व एक पल्याण (पुढील दिशेत बाक असलेला) असत (उदा., मायमोसेरस, ॲनार्सेस्टिस व अगोनियाटाइट) अगोनियाटाइटात मात्र खंड व पल्याण बोथट टोकाचे असतात. मायमोसेरस अपरिवर्तनशील होते. ॲनार्सेस्टिस व अगोनियाटाइट संजातींपासून आरारवैज्ञानिक (शारीरिक) वैशिष्ट्यांच्या विविध संयोगांद्वारे पुराजीव काळातील सर्व ॲमोनाइट उदयास आले. कवचाच्या घट्ट कुंडलनाने नाभी लहान होत गेली खंडांची संख्या वाढून खंड व पल्याण अधिक खोल होत गेले मंडलाचा (वलयाचा) छेद वर्तुळाकाराचा विवृत्ताकार ते अधर बाजूला निमूळता होत गेला व यामुळे डेव्होनियनच्या मध्यासच ॲमोनॉइड प्राण्यांत पुष्कळ विविधता आली होती. आर्सेस्टिस ग्लोयफिओसेरस अगोनियाटाइट प्रोलेकॅनाइट हे डेव्होनियनमधील अधिक महत्त्वाचे वंश होत. मेडलिकॉटिया, गोनियाटाइट, सायक्लोलोबस, पोपॅनोसेरस, टार्नाइट, ॲगाथिसेरस हे कार्बॉनिफेरस व पर्मियन काळांत अवतरलेल्यांपैकी महत्त्वाचे वंश होते. उत्तर पुराजीव काळात हे ॲमोनॉइड विपुल झाले व त्यांच्या विविध प्रकारांमुळे ते काळाच्या प्राणिजीतींपैकी प्रमुख गट होते.
या सर्वांमध्ये व वस्तुतः बहुसंख्य अमोनॉइडांमध्ये नवीन खंड व पल्याण मंडलाच्या नाभीकडील (पृष्ठीन) कडेजवळ निर्माण झाले व अशा तऱ्हेने सर्वांत जुने व मोठे खंड व पल्याण अधर बाजूच्या जवळ असतात. तथापि जेफ्युरोसेरस–बेलोसेरस शाखेत नवीन खंड व पल्याण नाभीच्या तसेच मुखीन कडेच्याही जवळ निर्माण होतात व अशा प्रकारे सर्वांत जुने व मोठे घटक मंडलाच्या मध्य बाजूला असतात. अशा तऱ्हेने ही सीता बहुसंख्य ॲमोनॉइडांतील सीतांच्या तऱ्हेहून अतिशय वैशिष्ट्यपूर्ण तऱ्हेने वेगळी आहे. केवळ उत्तर डेव्होनियनमध्ये आढळणाऱ्या क्लायमेनिइडी कुलातील प्राण्याचे कवच फासळीदार, सामान्यतः अधर बाजूस अरुंद व निमूळते आणि बहुतेक ॲमोनाइडांमधल्याप्रमाणे मध्य स्थितीपासून मुखीन बाजूकडे जाण्याऐवजी निनालिका पृष्ठीन बाजूला असते. हयातीच्या या सुस्पष्ट आणि थोड्या कालावधीमुळे हे कुल स्तरवैज्ञानिक सहसंबंधनिश्चितीमध्ये फार उपयुक्त आहे.
पुराजीव काळातील बहुतेक ॲमोनॉइड पर्मियनच्या अखेरीस निर्वंश झाले फक्त पुढील तीन वंशशाखा व संजाती ट्रायसिकमध्ये टिकून राहिल्या : (१) प्रोलेकॅनाइट : हा अपरिवर्तनशील गट होय, (२) जेफ्युरोसेरस : हा गट प्रगतशील असून त्याची सीता-तर्हाल वैशिष्टपूर्ण होती आणि (३) ग्लायफिओसेरस : हा गट खूप जोमदार ठरला व त्यापासून संपूर्ण ट्रायसिक काळामध्ये विविध वंशशाखा उदयास आल्या. जेफ्युरोसेरस-बेलोसेरस : या शाखेची प्रवृत्ती कवचेचे अधर बाजूला निमुळती असणे आणि झालरयुक्त अनेक खंड व पल्याण निर्मिणे यांकडे होती. या शाखेचा परामोत्कर्ष उत्तर ट्रायसिक काळातील पिनॅकोसेरस या वंशात झालेला दिसतो. प्रोलेकॅनाइट : या संजातीपासून मुख्यत्वे तीन वंशशाखा निर्माण झाल्या. सेराटाइट-टोयरोलाइट-शास्टाइट गटातील प्राण्याचे कवच खडबडीतपणे फासळीदार व त्याचे मंडल बाजूने कमीअधिक चापट झालेले आणि पुष्कळ खंड व पल्याणामध्ये पल्याण पूर्णतया किंवा कधीकधी अल्प प्रमाणातच भेगा असलेले व खंड दंतुरित असतात. एकूण हा गट अपरिवर्तनशील होता. लेकॅनाइट-जिम्नाइट गटातील कवचे सामान्यतः नितळ होती. सीतांच्या तळभागामुळे खंड व पल्याण यांचे विभाजन झालेले होते. ट्रॅचिसेरस हा तिसर्याव शाखेचा नमुनेदार प्रतिनिधी असून या शाखेत सीतांच्या तळभागामुळे जटिलता निर्माण झाली होती कवचांवर नक्षी होती व मंडल-छेद विविध प्रकारचा होता. अशा प्रकारे हा गट सेराटाइट व लेकॅनाइट गटांना तुलनेने अधिक प्रगतशील होता.
ग्लायफिओसेरस : ही संजाती प्रोलेकॅनाइटपेक्षा अतिशय जोमदार व प्रगत होती आणि तिच्यापासून प्रोलेकॅनाइटच्या शाखांना समांतर तीन वंशशाखा निर्माण झाल्या होत्या. अशा तऱ्हेने टिकायटीजची कवचे कमी प्रमाणात फासळीदार होती पण त्यांच्यातील सीतांची तऱ्हा जटील होती, ते मध्य व उत्तर ट्रायसिक काळाचे वैशिष्ट्य असून ते लेकॅनाइट-जिम्नाइट गटाशी तुल्य होत. ट्रॉपाइट हेही मध्य व उत्तर ट्रायासिक काळातील वैशिष्ट्यपूर्ण प्राणी आहेत. त्यांची कवचे अतिशय विविधतापूर्ण असून त्यांच्यामधील सीतांची जटील तर्हाह ही ॲमोनाइटातील वैशिष्टपूर्ण तऱ्हा आहे. त्यामुळे ते प्रोलेकॅनाइट संजातीमधील ट्रॅचिसेरस गटाशी तुल्य आहेत. आर्सेस्टिस शाखेतील कवचाचे सेराटाइटप्रमाणे कमी नक्षीदार होती. व त्यांच्यातील सीत जरी जटील होत्या, तरी त्या ट्रॉपाइट व टिकयटीज गटांपेक्षा साध्या होत्या.

ट्रायासिकमध्ये बहुसंख्य ॲमोनॉइडियांची अशी निरनिराळ्या दिशांनी भरभराट होत असताना काही अपरिवर्तनशील संजाती आणि प्रत्यावर्तन दर्शवीणाऱ्या शाखाही होत्या उदा., कोरिस्टोसेरस, काँक्लोसेरस, ऱ्हॅब्डोसेरस, पेरिप्ल्युराइट इत्यादींमधील विकुंडलनाची प्रवृत्ती अशा सर्व प्रवृत्ती ज्यात आढळतात तो ट्रयासिक कल्प ‘अमोनॉइडियांचे युग’ मानतात.
ट्रायासिक कळातील ॲमोनॉइडियांची बहुतेक कुले नष्ट झाली व ट्रायासिकनंतरचे ॲमोनाइडिया हे प्रत्यक्ष वा अप्रत्यक्षपणे मुख्यतः मोनोफायलाइट व सिंबाइट-नॅनाइट या संजातींपासून निर्माण झाले.
मोनोफायलाइट संजातीपासून जुरासिक काळात उदयास आलेली अधिक महत्त्वाची कुले म्हणजे सायलोसेरॅटिडी, लायटोसेरॅटिडी आणि फायलोसेरॅटिडी ही होत. सायलोसेरॅटिडी कुलातील प्राण्यांच्या कवचावरील सीतांची तऱ्हा मोनोफायलाइटसारखी असून अधर बाजू गोलसर असलेला सायलोसेरस व चापट बाजूला असलेला श्लोथायमिया या वंशांत कवचावर फासळ्या व घड्या यांची नक्षी निर्माण झालेली आढळते. ते फक्त पूर्व जुरासिकमध्येच आढळतात. लायटोसेरॅटिडी कुलातील सीतांची तऱ्हाही मोनोफायलाइटसारखी असते पण पल्याण आणि खंड साधे होताना दिसतात कवचावर बारीक फासळ्यांची नक्षी होती व बदलत्या पण कमी तऱ्हेने नाभी अधिकाधिक मोठी होत गेली. जुरासिकमध्ये व विशेषकरून पूर्व व मध्य क्रिटेशस काळांत पुष्कळ अकुंडलित वंश या कुलापासून उदयास आल्याचे मानतात आणि मॅकोस्कॅफाइट, स्कॅफाइट, क्रायोसेरस, हॅमाइट, टरिलाइट व बॅक्युलाइट हे त्यांपैकी अधिक लक्षात ठेवण्यासारखे वंश होत. फायलोसेरॅटिडी कुलात द्विचषकी पल्याण विकसित झाले. या कुलात पुष्कळ पल्याण व खंड होते अस्पष्ट फासळ्या किंवा पुसट घड्या व लहान नाभी होती. हे कुल क्रिटेशसच्या अखेरीपर्यंत टिकून होते व ते अपरिवर्तनशील असल्याने त्यात अत्यल्पच बदल झालेला आढळतो.
बहुतकरून नॅनाइटसारख्या संजातीपासून निर्माण झालेले ट्रायासिक नंतरचे इतर ॲमोनॉइडिया जगण्यासाठी पिरिस्थितीशी अतिशय दक्षपणे जुळवून घेण्याचा सतत प्रयत्न करीत होते व त्यांनी एकापाठोपाठ एक अशा तऱ्हेने अनेक कुलांना जन्म दिला. या प्रत्येक कुलाचे आयुष्य अल्प होते व या थोडक्या कालावधीत त्यांचा विस्तृत प्रसार झाला. यामुले जुरासिक व क्रिटेशस काळांचे अल्प कालावधीचे व विशिष्ट प्राणिसमूह असलेले उपविभाग पाडण्याच्या दृष्टीने ही कुले सुयोग्य ठरली आणि त्यामुळे विविध द्रोणींमधील स्तरांचे निकटचे सहसंबंध ठरविणे शक्य झाले. त्यांच्यामधील विविधता ही त्यांच्यातील पुढील घटकांच्या कित्येक संयोगांमुळे आली होती. कवचावरील नक्षीचे प्रकार, मंडल-छेद, सीतांचे पल्याण व खंड यांचे निरनिराळ्या अनुक्रमाने होणारे विभाजन व त्यापासून बनलेले विविध प्रकार अशा अनेक घटकांच्या संयोगामुळे या काळातील ॲमोनॉइडियांमध्ये इतकी विविधता आली होती.
या गटांचे पूर्व लायसमधील (जुरासिक कल्पाच्या सर्वांत आधीच्या विभागातील) अधिक महत्त्वाचे कुल एरिटिडी हे असून यातील परिवलयी ते चौकोनी प्रकारचे मंडल बदलून अरुंद वा अधर बाजूस निमुळते होत गेलेले होते. याच्यानंतर एकोसेरॅटिडी, डेरोसेरॅटिडी व पॉलिमॉर्फिडी ही कुले उदयास आली त्यांच्या कवचावरील फासळ्यांची वाढ आणि मंडल-छेदाचा उठावदारपणा ही सीतांच्या तऱ्हांपेक्षा अधिक ठळक होती. मध्य लायसमध्ये गोलसर मंडलाचे लिपारोसेरॅटिडी व तदनंतर दोरीसारखा विविष्ट कणा आणि कवचावर बळकट फासळ्या असलेले अमल्थिइडी हे कुल अवतरले.
हिल्डोसेरॅटिडी हा मध्य जुरासिक काळातील ॲमोनॉइडीयांचा प्रमुख गड होता. त्यांच्चा कवचावरील फासळ्या दात्राकार (चंद्रकोरीच्या आकाराच्या), कधीकधी मंडलाच्या मध्य बाजूवर छोटासा खळगा व अधर बाजूस कणा खोबण किंवा दोन्ही बाजूंना खोबणी असलेला कणा असे मंडल-छेद बहुधा जवळजवळ सपाट किंवा किंचित बहिर्वक्राकार होता. मध्य व उत्तर जुरासिक काळामध्ये स्फेरोसेरस, कॅडोसेरस व पेरिस्फिंक्टिस आणि त्यांच्याशी संबंधित असलेले ॲमोनॉइडीया सर्व सामान्य होते त्यांच्या कालावधी पूर्ण जुरासिक कालीन ॲमोनॉइडियाच्या कालावधीपेक्षा काहीसा जास्त होता. स्फेरोसेरस व कॅडोसेरस गटांतील प्राण्यांची कवचे अधर बाजूस रुंद आणि नाभी खोल व लहान होती. पेरिस्फिंक्टिस गटामधील कवचांची नाभी उथळ, मोठी होती व त्यावरील फासळ्यांचे अधर बाजूस फुटून दोन वा अधिक भाग झालेले होते तसेच मधूनमधून पुष्कळदा कवचांचे संकोचन झालेले आढळते.
होप्लिटिडी व ऑल्कोस्टेफॅनिडी ही पूर्व क्रिटेशस काळातील प्रमुख कुले होती. ऑल्कोस्टेफॅनिडी कुलामध्ये मंडल गोलसर, फासळ्या बळकट व नाभी रुंद होती. होप्लिटिडी कुलात कवचे विविध प्रकारची होती परंतु सर्वांच्या फासळ्या बळकट, साध्या वा दुभागलेल्या असून पुष्कळदा त्यांच्यावर गुलिकांच्या एक वा दोन ओळी असत बाजू किंचित बहिर्गोल आणि अधर बाजूवर बहुधा खोबणी, पण कधीकधी कणा असे.
ॲकँथोसोरॅटिडी हा मध्य क्रिटेशसमधील अतिशय प्रसार पावलेला व प्रमुख गट होता. यातील कवचावर अनेक बळकट फासळ्या व गुलिकांच्या कित्येक ओळी होत्या आणि पुष्कळदा मंडलाचा आडवा छेद चौकोनी व नाभी लहान होती. लायटोसेरॅटिडी कुलापासून आलेले अंकुडलित ॲमोनॉइडियाही मध्य क्रटेशसमध्ये सर्वसामान्यपणे होते. हे निरनिराळे अल्पकालीन ॲमोनॉइडिया गट होते, त्याच काळात बहेतेक सर्व क्रिटेशस काळभर आढळणारे डेस्मोसेरॅटिडी कुल अस्तित्वात होते. त्यांच्यातील कवचे मोठी, अधर बाजूस गोलसर व कमी नक्षीकाम असलेली होती. प्लॅसेंटिसेरॅटिडी हे जवळजवळ सर्व पूर्व व उत्तर क्रिटेशस काळात आढळणारे दुसरे महत्त्वाचे कुल असून त्यातील कवचाच्या बाजू किंचित बहिर्गोल, मंडल-छेद अधर बाजूस निमूळता होत जाणारा, अनेक जटिल खंड व पल्याण आणि पुष्कळदा पुसट व बुटक्या घड्यांची अस्पष्ट नक्षी त्यावर आढळते.
क्रिटेशसमध्ये ॲमोनाइडियांच्या कित्येक शाखा विविध दिशांनी विकसित होत असताना कमीअधिक प्रमाणात निगडित असलेल्या पुष्कळ संजातींमध्ये झालरी नष्ट होऊन व विभाजक रेषा अस्पष्ट होत जाऊन सीतांची तऱ्हा साधी होण्याकडे प्रवृत्ती होती. परिणामी या सर्व ॲमोनॉइडियांचे रूप ट्रायासिक काळातील सेरॅटिटॉइड या अपरिवर्तनशील गटातील शंखाप्रमाणे एकसारखे होत गेले. त्यांच्या एकत्रित गटाला स्यूडो-सेरॅटिटॉइड असे म्हणतात.

अशा प्रकारे जुरासिक व क्रिटेशस काळांत विपुल व विविध प्रकारचे ॲमोनॉइडिया होते. ते जेथे साचत त्या द्रोणींदधील परिस्थितीत जलदपणे बदल होत गेल्याचे पुढील गोष्टींवरून दिसते. अल्पजीवी कुले व एकापाठोपाठ आलेले गट हे झपाट्याने बदलत्या परिस्थितीशी जुळवून घेण्याचे प्रयत्न करित होते, हे निश्चित आहे. कवचाच्या विकुंडलनाची किंवा कमी घट्ट कुंडलनाची प्रवृत्ती आणि स्यूडो-सेरॅटिटॉइडांमध्ये ठळकपणे दिसून येणारी सीतांच्या तऱ्हा साध्या होत जाण्याची प्रवृत्ती यांच्यावरूनही परिस्थितीत सतत व झपाट्याने बदल होत गेल्याचे दिसून येते.
डेव्होनियनमध्ये (किंवा सिल्युरियनच्या शेवटी) अवतरल्यापासूनचा ॲमोनॉइडियांचा दीर्घकालीन इतिहास समृद्ध व विविधतापूर्ण आहे. तथापि क्रिटेशसच्या अखेरच्या काळात ते जलदपणे ऱ्हास पावत गेले आणि क्रिटेशसच्या अखेरीस निर्वंश झाले. त्यामुळे नवजीवन महाकल्पात पदार्पण करणारा त्यांचा वंशज शिल्लक राहिला नाही. [→ ॲमोनॉइडिया].
बेलेम्नॉइडिया : (सेफॅलोपाडा मॉलस्का). ऑक्टोपस, लोलिगो, सेपिया, स्पायरूला हे टेट्राब्रँकिएट सेफॅलोपाडा (शीर्षपाद) प्राण्यांचे हल्लीचे प्रतिनिधी असून जीवश्मांच्या दृष्टीने फक्त बेलेम्नॉइडिया गट महत्त्वाचा आहे. बेलेम्नॉइडियांच्या बाबतीत पश्चकवच (गार्ड) हा गजासारखा घटक भागच बहुतेक वेळा जीवश्मरूपात आढळतो. हे पश्चकवच अक्षाभोवतीच्या अरीय मांडणीच्या तंतुमय कॅल्साइटाच्या समाक्ष थरांचे बनलेले असते. याची पश्चबाजू निमुळती असते व त्याच्या अग्रस्थ टोकाजवळ शंकूच्या आकाराची पोकळी असते, तिला गर्त म्हणतात. तिच्यात आवृत्ती शंकू म्हणजे प्राण्याचे कोटरयुक्त कवच असते.
ट्रायासिकमध्ये किंवा पर्मियनअखेरीस ऑर्थोसेरॅटन संजातीपासून उत्पन्न झालेले ॲट्रॅक्टाइट आणि ओलॅकोसेरस हे वंश ट्रायासिक काळातील बेलेम्नाइटांचे प्रतिनिधी असून केवळ लांब आवृतशंकूच्या आदिकवचाचे म्हणजे प्राक्शंखाचे रक्षण करणारे छोटे पश्चकवच त्यांना होते. एका वंशशाखेत पश्चकवच जलदपणे वाढून जवळजवळ संपूर्ण कठीण संरचना त्याचीच बनली, तर लहान झाला. स्थूलमानाने यालाच बेलेम्नाइट म्हटले जाते. जुरासिक व क्रिटेशस काळांत याचा विस्तृत प्रसार झाला होता व स्थानिक किंवा मर्यादित भागांत त्यांची इतकी दाटी झालेली आढळते की, त्यावरून तेथिल निक्षेपाला बेलेम्नाइट शेल, बेलेम्नाइट मार्ल इ. नावे पडली आहेत. ते क्रिटेशसअखेर निर्वंश झाले.
बेलेम्नॉइडियाच्या नवजीव महाकल्पातील बेलोसेपिया व स्पायरूलिरोस्ट्रा या शाखांतील पश्चकवच हे आदिकवच आच्छादण्याइतके लहान झाले होते. ते जीवाश्मांच्या दृष्टीने महत्त्वाचे नाही. ऑक्टोपसच्या शाखेमध्ये कवच किंवा पश्चकवच निर्माण झाले नाही म्हणून त्याच्या पूर्वेतिहासाचा पुरावा जीवाश्मरूपात टिकून राहिला नाही. [→ बेलेम्नाइट].
फोरॅमिनीफेरा : (प्रोटोझोआ-जिम्नोमाक्सा किंवा सार्कोडिना). बहुतेक फोरॅमिनीफेरा सागरात राहणारे आहेत. बहुसंख्य नितलस्थ असून पुष्कळ तलप्लावी प प्लवकजीव आहेत, ते मुख्यत्वे उष्ण व उपोष्ण कटिबंधातील राहणारे असून त्यांचा प्रसार मोठ्या प्रमाणात तापमान व पाण्याची खोली यांवर अवलंबून असतो. बहुसंख्य फोरॅमिनीफेरांना चोल (कवच) असतो. चोल कायटिनमय, वालुकामय, सिलिकामय किंवा चूर्णीय असतो. कायटिनमय चोल जीवाश्मरूपात टिकून राहत नाही. सभोवतालच्या वाळूचे कण अथवा फुटलेल्या शंखशिंपाचे बारीक कण विविक्षित तऱ्हेने चिकटविले जाऊन रचिलेले असे वालुकामय चोल बनलेले असतात. स्रवणाद्वारे बनलेला चूर्णीय चोल सर्वांत सामान्यपणे आढळणारा असून सिलिसिनिडी कुलातील, तसेच मचूळ पाण्यात किंवा समुद्रात खोल जागी राहणाऱ्या मिलिओलिडी कुलातील काही प्राण्यांचे सिलिकामय चोल स्रवणाने बनलेले असतात. अध्ययनाच्या सोयीसाठी फोरॅमिनीफेरांचे पुढील गट करता येतात.
(१) लॅगिनॉइड गट : या गटातील प्राण्यांच्या भित्तीमध्ये सूक्ष्म छिद्रे असतात व परावर्तित प्रकाशात पृष्ठभाग काचेसारखा दिसतो. लॅगिना हा एकपुटक वंश असून त्याचा चोल फुग्यासारखा किंवा नासपतीच्या आकाराचा असतो. अनेकपुटकी वंशातील पुटक किंवा कोटर नोडोसारिया वंशाप्रमाणे सरळपणे एकामागून एक निर्माण होत जातात किंवा किस्टलॅरियाप्रमाणे सपाट कुंडलाच्या रूपात निर्माण होतात अथवा रोटॅलियाप्रमाणे मळसूत्रजरूपात निर्माण होत जातात व त्यामुळे चक्राभ चोल बनतो व या चोलात आधीची सर्व कोटरे पृष्ठीन बाजूने दिसतात, तर फक्त शेवटचे मंडल मुखीन बाजूचे दिसते. हे सर्व प्रकार सूक्ष्म असून खडक निर्माण करणारे म्हणून ते महत्त्वाचे नाहीत. याला काही अपवाद आहेत. त्यांच्या विपुलतेमुळे हल्लीच्या खोल समुद्रातील ग्लॉबिजेरीना ऊझांसारखे निक्षेप बनले आहेत. उदा., उत्तर अमेरिकेतील उत्तर पुराजीव काळातील एंडोथायरा चुनखडक आणि उत्तर अमेरिका, रशिया, जपान, चीन इ. प्रदेशांतील उत्तर कार्बॉनिफेरस काळातील फ्युस्यूलिना चुनखडक.
फोरॅमिनीफेरा कँब्रियनमध्ये अवतरले परंतु काही अपवाद वगळता त्यांच्या कार्बॉनिफेरस काळापर्यंत फारसा प्रसार झाला नव्हता. ट्रायासिकपासून ते विपुल होत गेले. पुराजीव काळात काचमय व वालुकामय प्रकारचे फोरॅमिनीफेरा हे मुख्य होत पण ज्यांना मोठे फोरॅमिनीफेरा म्हणतात त्यांची सुरुवात टेथिस समुद्रातील क्रिटेशस काळातील ऑपरक्युलिनासारख्या प्रकारापासून झाली व त्यांना जलदपणे महत्त्वपूर्ण स्थान प्राप्त झाले. त्यांचा विकास पुढील दोन शाखांत झाला : (न्युम्युलाइट ज्यापासून आले तो गट व (२) ऑर्बिटॉइड ज्यापासून आले तो गट.
ऑपरक्युलिना–न्युम्युलाइट–ॲसिलिना गटात चोल सपाट कुंडलित रूपात विकसित होऊन त्यामुळे आधीची मंडले प्रथम कमीजास्त प्रमाणात व अखेरीस पूर्णपणे झाकली गेली. यातील पटले दुहेरी भित्तीची असून दोन्ही पत्रांच्या मध्ये नाल होता आणि स्तंभ या चूर्णी संरचनेतून नाल भागशः आरपार गेलेले नव्हते. हे स्तंभ बहुधा पृष्ठभागी सूक्ष्म गुलिकांच्या रूपात दिसतात आणि ते पृष्ठभागावरील नक्षीकामाचा एक घटक होतात. हे चोल बिंबाभ असतात. [→ फोरॅमिनीफेरा].
फ्युस्युलिना, श्वागेरिना इत्यादीमधील कोटरे न्युम्युलाइट गटापेक्षा पुष्कळ रुंद असून आधीची बहुतेक कोटरे झाकलेली असून त्यामुळे तर्करूप (चातीच्या आकाराचा) चोल तयार होतो.
ऑपरक्युलिना–श्वागेरिना–हेटेरोस्टेगिना–सायक्लोक्लपियस शाखेमध्ये कोटरांचे विविध आकारांच्या कोटरिकांमध्ये विभाजन झालेले असते. कोटरांची उंची परिधीय दृष्ट्या वाढत असतानाही त्यांचे कोटरिकांमध्ये विभाजन होते. त्यामुळे अधिक मोठ्या कोटरिकांच्या मध्य-चकतीच्या दोन्ही बाजूंना लहान कोटरिकांच्या मालिका तयार होतात.
उत्तर क्रिटेशस व तृतीय कल्पात विशेषतः मायोसीनपर्यंत न्युम्युलाइट व ऑर्बिटॉइड गटांतील पुष्कळ मोठे फोरॅमिनीफेरा हे सूचक जीवाश्म आहेत व ज्या ठिकाणी खनिज तेलयुक्त थरांचे सहसंबंध ठरविणे अतिशय महत्त्वाचे आहे तेथील तेलाचा शोध घेण्याच्या दृष्टीने हे सूचक जीवाश्म म्हणून अतिशय महत्त्वाचे आहेत. न्युम्युलाइट, ॲसिलिना, ऑपरक्युलिना, ऑर्बिटॉइड, ऑर्बिटोलिना, लेपिडोसायक्लिना, ग्लोबोरोटॅलिया, ग्लोबोट्रंकाना, डिक्टिओकोनायडीज, डिस्कोसायक्लिना, मायोजिप्सिना हे मोठ्या फोरॅमिनीफेरांचे अधिक महत्त्वाचे काही वंश होत.
(२) मिलिओलिड गट : या गटातील फोरॅमिनीफेरांच्या भित्तींचे वैशिष्ट्य म्हणजे त्या अच्छिद्री (काही वंशांमध्ये आधीच्या अवस्थांत सच्छिद्र) असून त्यांचा पृष्ठभाग परावर्तित प्रकाशात पोर्सलीनसारखा दिसतो. कोटरे काहीशी रुंद आणि मिलीओला, ॲल्व्हिओलिना, ऑर्बिटोलाइट इत्यादींप्रमाणे आधीच्या कोटरांना जवळजवळ वेढणारी असतात. त्यामुळे एकूण चोल तर्करूप अथवा जाडसर बिंबाभ दिसतो. कार्नुस्पायरा या सपाट-सर्पिल कुंडलित नलिकेच्या टप्प्यापासून सुरुवात होऊन स्पायरोलॉक्युलिना, क्विंक्वेलॉक्युलिना, ट्रायलॉक्युलिना, बायलॉक्युलिना यांच्यामध्ये लांबट कोटरे थोड्या कालावधीने एकामागून एक निर्माण होत जातात. पेनेरोप्लिस-ऑर्बिक्युलिना-ऑर्बिटोलाइट या दुसऱ्या मालिकेत लॅगिना सजातींच्या ऑर्बिटॉइड गटाप्रमाणे कोटरिका निर्माण होतात, म्हणून या दोन शाखांच्या विकासाचे टप्पे सारखे आहेत. मिलिओलॉइड संजातीमधील प्राणीही सूचक जीवश्म म्हणून उपयुक्त असून मिलिओला, ऑर्बिटोलाइट, ॲल्व्हिओलीना हे खडक निर्माण करणाऱ्या (उदा., इओसीन काळातील ) वंशांपैकी अधिक महत्त्वाचे वंश होत.
(३) वालुकामय गट : क्वॉर्ट्झ, स्पंजांच्या कंटीका आणि इतर प्राण्यांच्या कवचांचे तुकडे यांचे कण एकत्रित आणले जाऊन व विशिष्ट प्रकारच्या रचना करून ते चिकटविले जाऊन, या गटातील फोरॅमिनीफेरांचे चोल बनतात. हे बाह्य कण निवडताना व त्यांची मांडणी करतानाही अनेक वेळा बरेच तारतम्य दाखविल्याचे दिसून येते. या कणांना एकमेकांशी जोडणारे संयोजक द्रव्य किंवा लुकण कायटिनमय, लोहमय, चूर्णीय अथवा सिलिकामय असते. उष्ण कटिबंधात उथळ पाण्यात राहणाऱ्या काही चोल प्रकारांत चूर्णीय लुकण असते, तर मचूळ व खोल पाण्यातील काही प्रकारांत (उदा., सिलिसिनिडी कुलातील) सिलिकामय लुकण असते.
जे. ए. कुशमन यांच्या विचारप्रणालीनुसार वालुकामय चोल आदिम (प्रिमिटिव्ह) वैशिष्ट्य आहे कारण हे प्रकार स्रवणाद्वारे चोल बनविण्यास असमर्थ होते, असे मानतात परंतु जे. जे. गॅलोवे यांच्या मतप्रणालीनुसार सॅकमिनिडी, ॲस्ट्रोऱ्हायझिडी इत्यादींसारख्या काही कुलांचे वालुकामय चोल हे आदिम वैशिष्ट्य असतेच. शिवाय पर्याजवरणाशी जुळवून घेण्यासाठी किंवा कवच स्रवण्याची क्षमता ऱ्हास पावली वा कमी झाली म्हणून प्रगत कुलांमध्येही वालुकामय चोल निर्माण झाल्याचे आढळते. अवशेष रूपात टिकून राहणाच्या दृष्टीने वालुकामय चोल अयोग्य असल्याने अधिक जुन्या शैलसमूहांमध्ये ते विरळाच आढळतात.
काही वालुकामय फोरॅमिनीफेरांमधील चोलांची संरचना ही लॅगिना संजातीमधील संरचनेसारखी असते. उदा., सॅकमिनातील लॅगिनासारखी, रिओफॅक्समधील नोडोसारियासारखी व ट्रोकॅमिनातील रोटॅलियासारखी असते. [→ फोरॅमिनीफेरा].
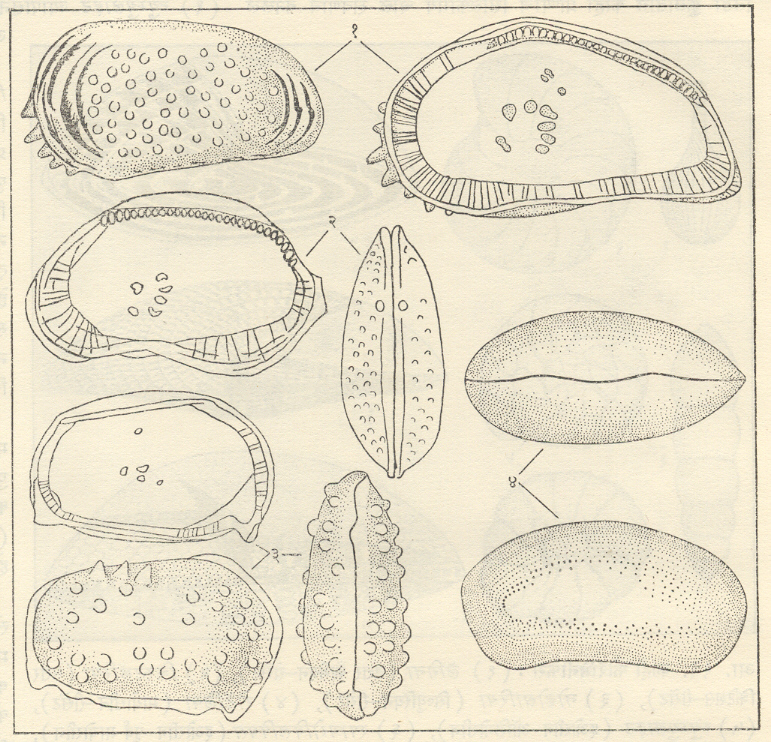
ऑस्ट्रॅकॉडा : (क्रस्टेशिया, आर्थ्रोपोडा). हे सूक्ष्म प्राणी पृष्ठवर्मात (ढालीसारख्या संरचनेत ) झाकले गेलेले असतात. पृष्ठवर्माची दोन शकले असून ती पृष्ठीन बाजूला लवचिक बंधाने किंवा दात व कटक यांच्या अंतर्बंधनाने एकमेकांना जोडलेली असतात. या शकलांची हालचाल त्यांच्या मध्यालगत अग्र बाजूला असलेल्या अभिवर्तनी स्नायूंद्वारे होते. पृष्ठवर्म चूर्णीय, फॉस्फेटी किंवा अचूर्णीय कायटिनमय द्रव्याचे असू शकते. ते सामान्यतः १ मिमी. पेक्षा कमी व क्वचित ५ मिमी.पर्यंत लांब असते. त्याची बाहेरची बाजू गुळगुळीत वा विविध प्रकारचे नक्षीकाम असलेली असते. ऑस्ट्रॅकॉडांच्या बहुतेक जीवाश्मांमध्ये बिजागरीची रचना, नक्षीकाम, अभिवर्तनी स्नायूंचे ठसे व कधीकधी ज्यांमुळे उपांगांची हालचाल होते त्या स्नायूंचे ठसे टिकून राहिलेले आढळतात.
बहुतेक ऑस्ट्रॅकोडा जलचर असून त्यांचे जवळजवळ सर्व प्रकारच्या जलीय परिस्थितींशी अनुकूलन झालेले आढळते उदा., नदी, गोड्या पाण्याचे सरोवर व डबकी, तसेच समुद्र, नदीमुख, खारे सरोवर, पाणथळ भाग, शिवाय गंधकयुक्त झरे यांसारख्या विविध प्रकारची लवणता असलेल्या जलीय स्थितींत ते राहतात. काही जमिनीवरही राहणारे आहेत उदा., मेसोसायप्रिस टेरेस्ट्रिॲलिस (आफ्रिकेतील क्निस्नवन). हे बहुतेक रांगणारे वा बीळ करून राहणारे नितलस्थ प्राणी असून काही तलप्लावी आहेत, तर काही समुद्राच्या तळाजवळ व तेथील वनस्पतींमधून थोडे अंतर पोहून जाणारे आहेत. सूक्ष्मकणी गाळ व विपुल जैव अन्न असलेल्या भागांत हे सर्वसामान्यपणे आढळतात आणि भरड वाळू व जोरदारपणाने वाहणाऱ्या नद्यांसारख्या अस्थिर स्थितींमध्ये ते विरळच आढळतात. नदीमुखीय परिस्थितींमध्ये ते फोरॅमिनीफेरांच्या संदर्भात पूरक असे प्राणी असून अशा प्रकारे त्यांच्या साह्याने समुद्रकिनाऱ्याचे स्थान दाखविता येते आणि त्यामुळे समुद्राचे आक्रमण व माघार यांची व्याप्ती दाखविण्याच्या दृष्टीने ऑस्ट्रॅकॉड प्राणी उपयुक्त असतात. चूर्णीय फोरॅमिनीफेरा असलेल्या गाळात ते ऑस्ट्रॅकॉडा बहुधा नसतात आणि कोनोडांटा असलल्या गाळात ते आढळत नाहीत. विविध खोलींवर व भिन्नभिन्न अक्षांसामध्ये ऑस्ट्रॅकॉडांचे नियंत्रण बऱ्याच प्रमाणात तापमानाच्या मर्यादांद्वारे होत असते.
अशा प्रकारे निक्षेपणाच्या परिस्थितीशी ऑस्ट्रॅकॉडांचा निकटचा संबंध असतो आणि पर्यावरणाद्वारे नियंत्रित होणाऱ्या ऑस्ट्रॅकॉडांच्या स्वरूपामुळे खनिज तेलाच्या पूर्वेक्षणाच्या दृष्टीने ऑस्ट्रॅकॉडा हा प्राणीगट अतिशय महत्त्वाचा असल्याचे आढळून आले आहे. [→ आर्थ्रोपोडा].
कोनोडोंटा : कोनोडोंटा अवशेष म्हणजे साध्या अथवा जटिल सूक्ष्म संरचना असून त्यांचे आकारमान ०.१ मिमी. पेक्षा कमी व ४ मिमी. पेक्षाही जास्त असू शकते. रासायनिक व प्राकृतिक झीज होण्याला हे अवशेष काहीसे कणखर असून ते दुधी काचेसारखे पारभासी व अंबराप्रमाणे उदी रंगाचे असतात. वातावरणाची क्रिया झाल्यावर ते अपारदर्शक व करडे ते काळे होतात. त्यांचे माशांच्या दातांशी खूप साम्य असून दीर्घ काळापासून त्यांचा अभ्यास तसेच मानून झाला होता. वलयी (ॲनेलिडा) प्राण्यांच्या जबड्याचा भाग, आर्थ्रोपोडाच्या कंटिका, कृमीसारखा प्राण्यांच्या मैथुन-कंटिका, शैवालांनी स्रवलेल्या दंतुरित संरचना, गॅस्ट्रोपोडांचे रेत्रिकीय दात व मिसूरी येथील ऑर्डोव्हिसियन कालीन आर्किओग्नॅथस या संभाव्य माशाचे दात म्हणजे कोनोडोंटा होत, असेही मानले गेले होते परंतु माशांचे दात [सी. एच्. पँडर (१८५६), के. ए. त्सिटेल व जे. व्ही. रोहन (१८८६)] व वलयी प्राण्यांच्या जबड्यांचे भाग [एच्. डब्ल्यू. स्कॉट (१९३४) एच्. श्मिट (१९३४ आणि एफ्. एच्. टी. ऱ्होड (१९५४)] ही दोन स्पष्टीकरणे अधिक संभाव्य समजली जातात.

कोनोडोंटा ह्या अंतर्गत किंवा बाह्य सांगाड्याच्या संरचना आहेत हेही अजून प्रस्थापित झालेले नाही परंतु बहुधा त्या कोणत्या तरी ऊतकांच्या (समान रचना व कार्य असलेल्या कोशिकांच्या समूहांच्या) आत जडविलेल्या होत्या असे दिसते.
पुराजीव कालीन गाळात कोनोडोंटा विपुल असून ट्रायासिकमध्ये त्यांच्या ऱ्हासास सुरुवात झाली. क्रिटेशस काळानंतर ते आढळतात की नाही याविषयी निश्चित काही सांगता येत नाही. कोनाडोंटा चुनखडकांत विपुलपणे, शेलांमध्ये सामान्यपणे, पण गाळवटी खडकांत व वाळूत विरळपणे आढळतात. ते ज्या खडकांत आढळतात त्याचा प्रकार व त्यांच्याबरोबर आढळणारे इतर जीवाश्म यांच्यावरून कोनोडोंटा हे सागरी, बहुतकरून तलप्लावी प्राण्याच्या शरीराचे भाग असावेत असे दिसते. ज्या समुदायांत ते आढळतात त्यांपैकी काहींमधील महत्त्वाच्या घटकांतील असममितीवरून (असमानतेवरून)ते ज्या प्राण्यांशी निगडित होते त्यांपैकी काही प्राणी तरंगत्या निवाहांच्या स्वरूपात राहत असावेत असे दिसते.
कोनोडोंटा कोणत्या प्राणिगटाशी संबंधित होते व त्यांनी त्या प्राण्याच्या शरीरात कोणते कार्य केले याची आपल्याला माहिती नसली, तरी विशेषकरून पुराजीव कालीन थरांमधील मुख्यत्वे समुदायाच्या रूपातील त्यांच्या विपुलता ही जैव स्तरवैज्ञानिक कार्या ला उपयुक्त आहे. ते ज्या संभाव्य तलप्लावी पर्यावरणातून बुडून समुद्रतळावर गेले ते पर्यावरण कोणते असावे, हे या विपुलतेद्वारे सूचित होते. रासायनिक विद्राव व प्राकृतिक अपघर्षण (झीज) यांना विरोध करण्याच्या गुणांमुळे त्यांचे पुनरावर्तन झाले असण्याची शक्यताही ते ज्या गाळात असतात तो साचण्याची परिस्थिती स्पष्ट करताना नेहमी लक्षात ठेवली पाहिजे.
चुनखडक, सूक्ष्मकणी शेल व ग्रॅप्टोलाइटयुक्त काळे शेल अशांसारख्या खोल पाण्यात साचणाऱ्या खडकांत कोनोडोंटा सामान्यतः आढळतात, असे सूचित होते परंतु दगडी कोळशाचे थर असलेल्या चुनखडकांतही (उदा., इंग्लंडमधील कारबॉनिफेरसकालीन) ते आढळतात आणि त्यावरून ते उथळ पाण्यातही साचल्याचे दिसून येते. पायराइट, ग्लॉकोनाइट व फॉस्फोराइट ही खनिजे पुष्कळदा कोनोडोंटाबरोबर आढळतात व त्यांच्यावरून नितलस्थ प्राण्यांना प्रतिकूल अशी क्षपणकारी (ऑक्सिजन कमी प्रमाणात असणारी किंवा त्याचा अभाव असणारी) परिस्थिती त्या द्रोणीत असल्याचे सूचित होते.

मार्ग-चिन्ह द्रीवाश्म : जीवाश्मांच्या या गटात प्राण्यांच्या पावलांचे ठसे, वाटचालीच्या खुणा, बिळे, त्यांनी पाडलेली छिद्रे इ. येतात. प्राण्यांच्या कृतीचे प्रतीबिंब या जीवाश्मांत पडलेले दिसते. अगदी भिन्न गटांतील प्राण्यांकडून सारखे किंवा अगदी सारखे मार्ग-चिन्ह जीवाश्म निर्माण झालेले असू शकतात. त्यामुळे या गटातील जीवाश्मांचे अध्ययन करताना वापरलेल्या वंशवाचक व विशिष्ट नावाचा अर्थ वापरात असलेल्या जीवविज्ञानीय पद्धतीतील नावाप्रमाणेच असेल, असे नाही. त्यांचा उल्लेख रूपवंश व रूपजाती असा केला जातोच म्हणून यांचे मुख्य वर्गीकरण हे जगण्याच्या परिस्थिशी अनूरूप अशाच शब्दांत म्हणजे पावलांचे ठसे, वाटचालीच्या खुणा, बिळे व छिद्रे असेच असू शकते. मुख्यत्वे रूडोल्फ रिश्टेर यांनी सुरुवात केल्यावर ए. सैलाकेर व डब्लू. हेन्ट्शेल यांनी अतिशय काळजीपूर्वक अध्ययन करून (१) रेपिक्निया, (२) पॅसिक्निया, (३) फोडिक्निया, (४) डोमिक्निया व (५) क्युबिक्निया यांसारख्या अशा जीवाश्मांचा अर्थ लावणारी जीवपरिस्थितिवाज्ञानीय (आसमंतातील परिस्थितीनुरूप होणाऱ्या प्राण्यांच्या वर्तनाचा अभ्यास करणाऱ्या विज्ञानाची) परिभाषा निर्माण केली.
पावलांचे ठसे, प्राण्यांच्या वाटचालीच्या खुणा व पृष्ठभागालगतची क्षितिजसमांतर अथवा जवळजवळ क्षितिजसमांतर बिळे ही नितलस्थ व कधीकधी नितल-तरणक प्राण्यांनी विविध खोलींवर तयार केलेली असतात परंतु जेथे लाटा व प्रवाह यांचे विनाशक परिणाम होत नाहीत तेथे हे जीवाश्म सर्वसामान्यपणे टिकून राहिलेले असतात शिवाय अशा ठिकाणचा गाळ, पावलांचे ठसे, वाटचालीच्या खुणा व बिळे निर्माण होण्याइतपत गाळाचा थर सूक्ष्मकणी (मऊ) असावा लागतो व त्यात अन्नघटकही असावे लागतात, कारण त्याकरिता प्राणी येथे वरचेवर येतात. अशा प्रकारे हे ठसे (वा चिन्हे) अधिवेलीय व विक्षुब्ध आणि समुद्रतटीय पट्ट्यांमध्ये टिकण्यासारखे नसतात, मात्र जेथे या विनाशक घटकांचा परिणाम होत नाही अशा खंड–फळीच्या भागात हे जीवाश्म आढळतात. टणक खडक व घट्ट जाड गाळ यांच्यातील जवळजवळ उदग्र (उभ्या) बिळात (छिद्रात) प्राणी बहुधा कायमचे घर करून राहतात व सभोवतालच्या पाण्यात निलंबित (लोंबकळत्या) स्वरूपात असलेले अन्न त्यांना मिळते. कठीण खडकांतील किंवा घट्ट गाळातील छिद्र स्थानबद्ध प्राण्यांना आधार मिळावा म्हणून केलेली असतात. परजीवी (इतर जीवांवर उपजीविका करण्याच्या) जीवनप्रणालीसाठीही प्राण्यांच्या कवचामध्ये अशी छिद्रे पाडली जातात अथवा केवळ सहभोजी (दोन जीवांत एकमेकांना फायदेशीर ठरेल असे बाह्य साहचर्य असणाऱ्या) जीवनप्रणालीचा आधार म्हणून व आश्रयी प्राणी जेथे जेथे जातो तेथे तेथे अन्न मिळविण्याकरिता अशी छिद्रे निर्माण केलेली असतात.
म्हणून या जीवाश्मांचा अभ्यास ते निर्माण करणाऱ्या प्राण्यांची पर्यावरणीय परिस्थिती विशद करण्यासाठी करावा लागतो. स्तरवैज्ञानिक सहसंबंध ठरविणे किंवा शैलसमूहांची भूवैज्ञानिक वये काढणे यांसाठी हे जीवाश्म उपयुक्त ठरतील, अशी अपेक्षा नसते. हे मार्ग-चिन्हांचे अवशेष पाण्याच्या प्रवाहाने किंवा लाटांच्या माऱ्याने झिजून जाऊ शकतात परंतु शंखशिंपले किंवा इतर प्रकारची कवचे त्यांच्या मूळ स्थानापासून वाहून नेली जाऊन दुसरीकडे कोठे तरी साचू शकतात, तसे या मार्ग-चिन्हांच्या अवशेषांचे होऊ शकत नाही, हा महत्त्वाचा मुद्दा लक्षात घेतला पाहिजे. तथापि जर ज्यांमध्ये हे जीवाश्म आहेत असे दगडगोटेच वाहून नेले गेले असले, तर त्या दगडगोट्यांचे आणि त्यावरून त्यातील मार्गचिन्ह जीवाश्मांचे मूलस्थान माहीत होऊ शकते. [→ जीवाश्म].
अपृष्ठवंशी जीवाश्म गौण गट : रेडिओलॅरिया : (प्रोटोझोआजिम्नोमिक्सा किंवा सार्कोडिना). प्रोटोझोआतील रेडिओलॅरियागणाचे जीवाश्म एकूण प्राण्यांच्या जीवाश्मांमध्ये महत्त्वाचे नाहीत.
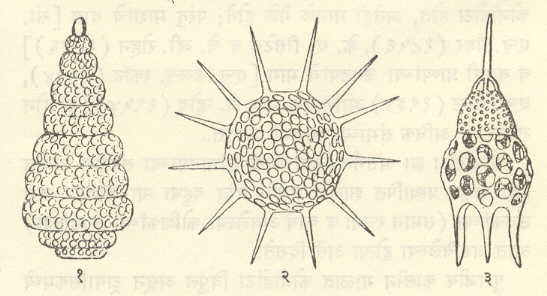
कँब्रियन-पूर्व काळातील त्यांच्या आढळाविषयी शंका आहेत मात्र कँब्रियन काळातील रेडिओलॅरियांचे जीवाश्म निश्चितपणे आढळले आहेत. या प्राण्यांचा सांगाडा सामान्यपणे कंटिकांचा किंवा विविध आकारांच्या नाजूक जालिकांचा बनलेला असतो. काहींचा सांगाडा स्ट्राँशियम सल्फेटाचा बनलेला असतो पण सामान्यपणे तो सिलिकेचा बनलेला असतो व तो जीवाश्मरूपात टिकून राहतो. सर्व रेडिओलॅरिया सागरात राहणारे असून बहुसंख्य तलप्लावी आहेत आणि ते ४०० मी. खोलीपर्यंत राहतात. प्राणी मेल्यावर हे सांगाडे सागरतळावर साचतात व रेडिओलॅरियन ऊझ नावाचे सूक्ष्मकणी सिलिकामय निक्षेप तयार होतात. उदा., हल्लीच्या हिंदी व पॅसिफिक महासागरांच्या उष्ण कटिबंधीय प्रदेशांतील अधिक खोल भागात आढळणारे रेडिओलॅरियन ऊझ. रेडिओलॅरियांच्या जीवाश्मांद्वारे पुष्कळदा उष्ण कटिबंधीय भागातील खोल सागरात निक्षेपण (साचण्याची क्रिया) झाल्याचे सूचित होते. [⟶रेडिओलॅरिया].
पोरिफेरा: (स्पंज). स्पंजाचे सांगाडे कंटिकाचे बनलेले असतात. कंटिका चूर्णीय वा सिलिकामय द्रव्याच्या बनलेल्या असू शकतात व त्यांवरुन स्पंजांचे (१) कॅल्केरिया (चूर्णीय) व (२) नॉन-कॅल्केरिया (अचूर्णीय) असे गट पडतात. पुष्कळदा हे सांगाडे दृढ नसतात व प्राण्यांच्या मृत्यूनंतर कंटिका सुट्या होतात. त्यामुळे असे जीवाश्म स्तरवैज्ञानिक दृष्टीने अतिशय निकामी व अल्प महत्त्वाचे ठरतात. मात्र जेव्हा एखाद्या ठिकाणी इतर जीवाश्म नसातात तेव्हा ते थोडेफार उपयुक्त असू शकतात तथापि काही संरचना टिकून राहतात. कॅल्केरिया गट ऑर्डोव्हिसियन काळात अवतरला आणि क्रिटेशस-जुरासिक काळातील स्पंजांचे चांगले प्रातिनिधिक वंश म्हणजे रॅफिडोनेमा व बॅरॉइसिया हे होत. नॉन-कॅल्केरियांपैकी लिथिस्टीड स्पंजांचे सांगाडे बळकट असल्याने ते चांगले टिकून राहिले असून ते उत्तर कँब्रियन काळात अस्तित्वात होते, ते जुरासिक व क्रिटेशस काळांत विपुल झाले होते व तेव्हाचे माहीत असलेले वंश पुढील होत: सायफोनिया, जॅरिया, व्हेरुक्युलिना, डोरिडर्मा. हेक्झॅक्टिनेलिडातील काही प्राणी पूर्व कँब्रियनमध्ये होते व हेक्झॅक्टिनेलिडही जुरासिक व क्रिटेशस काळांत विपुल झाले, त्यांपैकी व्हेंट्रिक्युलाइट हा वंश अस्तित्वात असल्याची नोंद आहे.
मोनॅक्सोनिडचा अपवाद वगळता स्पाँजिल्ला वंश व त्याच्याशी संबंधित असलेले सर्व स्पंज सागरात राहणारे असून सध्या ते जगाच्या सर्व भागांत आढळतात. हे स्पंज किनाऱ्यापासून सु.४०० मी. खोलीपर्यंतच्या भागात अधिक विपुल आहेत. [⟶ पोरिफेरा].
सीलेंटेरेटा: (हायड्रोझोआ व सिफोझोआ). हायड्रोझोआ वर्गातील स्ट्रोमॅटोपोरॉयडिया गणातील प्राण्यांचे सांगाडे चूर्णीय असून ते एककेंद्री पटलांचे बनलेले असतात. ते ओबडधोबड किंवा गोलाभ पुंजक्यांच्या आणि पुष्कळदा लेपांच्या (पापुद्रयांच्या) रूपात आढळतात. पुराजीव काळात या प्राण्यांपासून खडक बनल्याचे माहीत असून डेव्होनियन काळात ते विशेष विपुलपणे आढळतात. स्ट्रोमॅटोपोरा. अँफिपोरा, ॲक्टिनोस्ट्रोमाइ. मध्यजीव काळातही आढळतात.
शंखवासी (हर्मिट) खेकडा राहत असलेल्या विशेषकरून गॅस्ट्रोपोडांच्या कवचांभोवती लेपरूपात असलेले जिग्नोब्लास्टिया गणाचे म्हणता येतील, असे कायटिनमय सांगाडे आढळले आहेत. या गणाचे काही जीवाश्म यूरोपमधील ट्रायासिक, जुरासिक व पूर्व क्रिटेशस काळांतील खडकांत आढळले आहेत.
कॅलिप्टोब्लास्टिया गणाचे म्हणता येतील असे कायटिनमय सांगाडे व्हिक्टोरियामधील कँब्रियन कालीन व न्यूयॉर्क येथील ऑर्डोव्हिसियन काळातील खडकांत आढळले असून एक प्लाइस्टोसीन (सु. ६ लाख ते ११ हजार वर्षांपूर्वीच्या) काळातील निक्षेपातही सापडला आहे.
हायड्रोकोरॅलिना गणातील प्राण्यांचे सांगाडे चूर्णीय होते व ते दंडाचे (कांड्यांचे) जाळे बनून तयार झालेले असतात. ते पापुद्यांच्या किंवा शाखायुक्त स्वरूपाचे असतात. खडकनिर्मितीच्या दृष्टीने हे प्राणी महत्त्वाचे आहेत. हल्लीचा मिलेपोरिडियम हा वंश पोवळ्यांशी निगडित असून तो मायोसीन काळातही आढळला आहे. स्ट्रँबर्ग येथील उत्तर जुरासिक कालीन खडकांत मिलेपोरिडियम आणि ईजिप्तमधील क्रिटेशस कालीन खडकांत मिलेस्ट्रोमा हे वंश आढळले आहेत.
ज्यांमध्ये जेलीफिश येतात त्या सिफोझिआ प्राण्यांना कठीण अवयव नसले, तरी त्यांचे ठसे आढळतात उदा., बव्हेरियातील लिथोग्राफिक चुनखडकांतील ऱ्हायझोस्ट्रोमाइट व स्वीडनमधील कँब्रियन खडकांतील मेंडयुसिना. [⟶ सीलेंटेरेटा].
ॲनेलिडा: प्राचीन खडकांमधील बिळे, वाटचालीच्या खुणा ह्या कृमींमुळे निर्माण झाल्याचे कधीकधी मानतात परंतु त्यांच्या अनिश्चित स्वरुपांमुळे त्यांना पुष्कळदा मार्ग-चिन्ह जीवाश्म असे संबोधतात.
तथापि ॲनेलिडांपैकी पॉलिकीटांचे पुष्कळ जीवाश्म माहीत आहेत. हे जीवाश्म सरळ, वाकड्या, नागमोडी, सर्पिल वा इतर रीतीने कुंडलित अशा विविध प्रकारच्या नळ्यांच्या रूपात असतात. ते स्त्रवणाद्वारे बनविलेले (उदा.,कायटिनमय वा चूर्णीय) आणि बाह्यकण एकत्रित चिकटविले जाऊन बनलेले असू शकतात. त्यांच्या आढळाविषयी व प्रसाराबद्द्ल पुष्कळ माहिती उपलब्ध आहे. काही गोड्या व मचूळ पाण्यातही रहात असल्याचे माहीत असले, तरी ॲनेलिडा बहुतकरून सागरात राहणारे आहेत. ते उत्तर कँब्रियनमध्ये अवतरल्याचे निश्चितपणे माहीत आहे. ऑलिगोकिटा हा ॲनेलिडांचा दुसरा वर्ग कार्बॉनिफेरस काळापासून अगदी विरळाच आढळत असून शिवाय त्याचे जीवाश्म चांगले टिकून राहिलेले नाहीत.
कृमींपैकी सर्वांत चांगले माहीत असलेले जीवाश्म पॉलिकीटांचे आहेत, मात्र शैलसमूहांची भूवैज्ञानिक वये ठरविण्यासाठी खुद्द पॉलिकिटांचे जीवाश्म उपयुक्त असल्याचे अजून मानीत नाहीत परंतु जेव्हा इतर जीवाश्म उपलब्ध नसतात तेव्हा ते ज्यांत आढळतात त्या थरांचे भूवैज्ञानिक वय पॉलिकीटांच्या जीवाश्मांद्वारे स्थूलमानाने ठरविता येऊ शकते. [⟶ ॲनेलिडा].
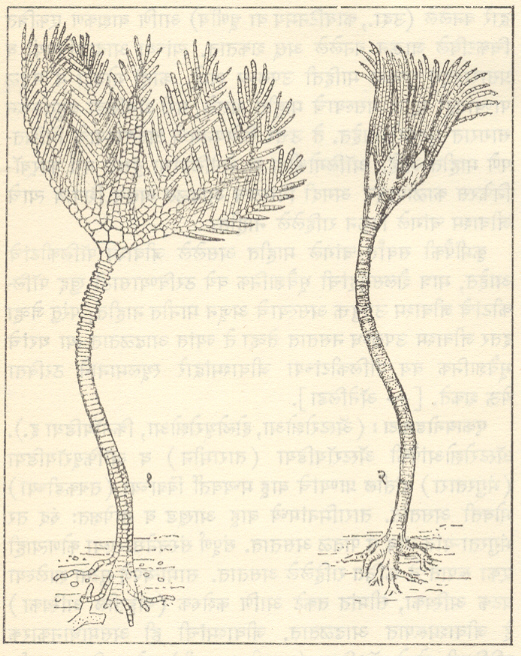
एकायनोडर्माटा: ( ॲस्टरोझोआ, होलोथूरोझोआ, क्रिनॉयडिया इ.). ॲस्टरोझोआंपैकी ॲस्टरॉयडिया (तारामीन) व ऑफियूरॉयडिया (भंगुरतारा) यांतील प्राण्यांचे बाहू मध्यवर्ती बिंबाच्या (तबकडीच्या) भोवती असतात. तारामिनांमध्ये बाहू आखूड व सापेक्षतः रुंद तर भंगुरता-यांत लांब व पातळ असतात. संपूर्ण संरचनेसारखा कोणत्याही एका रूपात ते क्वचित राहिलेले असतात. सामान्यपणे सुट्या झालेल्या घटक अस्थिका, सीमांत तकटे आणि कशेरुक (बाहूतील अस्थिका) हे जीवाश्मरूपात आढळतात. जीवाश्मांची ही असमाधानकारक परिस्थिती होलोथूरॉडिया (सागरी काकडी) मध्ये अधिकच वाईट झालेली दिसते. काळाइतक्या पूर्वीपासून आढळतात. ॲस्टरॉयडिया व ऑफियूरॉयडिया यांच्या हल्लीच्या प्रकारांशी अगदी सारखे असणारे प्रकार जुरासिक काळाइतक्या पूर्वीपासून आढळतात. ॲस्टरॉयडिया, ऑफियूरॉयडिया व होलोथूरॉयडिया कँब्रियन काळात अवतरले. हल्ली ते मध्यम खोल व बहुसंख्य ६० मी. पेक्षा कमी खोल पाण्यात राहत आहेत.
एकायनोडर्माटामधील पेल्मॅटोझोआंपैकी क्रिनॉयडिया (समुद्रनलिनी) सर्वांत महत्त्वाचे असून ते मुख्यत्वे पुराजीव कालीन आहेत. कँब्रियन ते पर्मियन काळातील सिस्टॉयडिया, ऑर्डोव्हिसियन ते पर्मीयन काळातील ब्लॅस्टॉयडिया व कँब्रियन ते कार्बॉनिफेरस काळातील एड्रिॲस्ट्ररॉयडिया यांच्या कटोरातील सममिती मूलतः पंचतयी असून बिंबाभतकटांनी बनलेल्या स्तंभाने कटोर समुद्रतळाला चिकटलेला असतो. क्रिनॉयडियामध्ये बाहू स्पष्ट असून सिस्टॉयडियामध्ये ते लहान झालेले असतात तर ब्लॅस्टॉयडिया व एड्रिॲस्टरॉयडिया यांच्यात ते नसतातच. उत्तर क्रिटेशसमधील यूईटॅकायनस व मार्सुपाईंट यांसारख्या काही क्रिनॉयडियामध्ये हा स्तंभ नसतो. ते तरंगणारे प्राणी असावेत. त्यांचा अल्पकालीन इतिहास व विस्तृत प्रसार यांमुळे ते उत्तर क्रिटेशस काळातील अतिशय उपयुक्त असे सूचक जीवाश्म ठरले आहेत. ट्रायासिकपासून क्रिनॉयडियांचा ऱ्हास होत असून आता त्यांचे सु. १०० वंश आहेत व ते बहुतेक उथळ पाण्यात राहत आहेत. पुराजीवमध्ये व विशेषतः कार्बॉनिफेरसमध्ये क्रिनॉयडिया हा एकायनोडर्माटांपैकी सर्वांत महत्त्वाचा गट होता. ॲक्टिनोक्रायनस, पोटेरिओक्रायनस, प्लॅटिक्रायनसव ऱ्होडोक्रायनस हे कार्बॉनिफेरसधील, तर पेंटॅक्रायनस, अँटेडॉनव मिलेरिक्रायनस हे जुरासिक काळातील अधिक महत्त्वाचे वंश होत. कार्बॉनिफेरस काळात ते इतके विपुल होते की, त्या काळात त्यांच्यापासून मोठ्या प्रमाणात क्रिनॉयडी चुनखडक व संगमरवर बनले होते. सिस्टॉयडिया, ब्लॅस्टॉयडिया व एड्रिॲस्टरॉयडिया हे गट त्यांच्या आयुष्यात नेहमीच गौण होते व त्यांचे जीवाश्म कमी प्रमाणात आढळतात. [⟶ एकायनोडर्माटा].
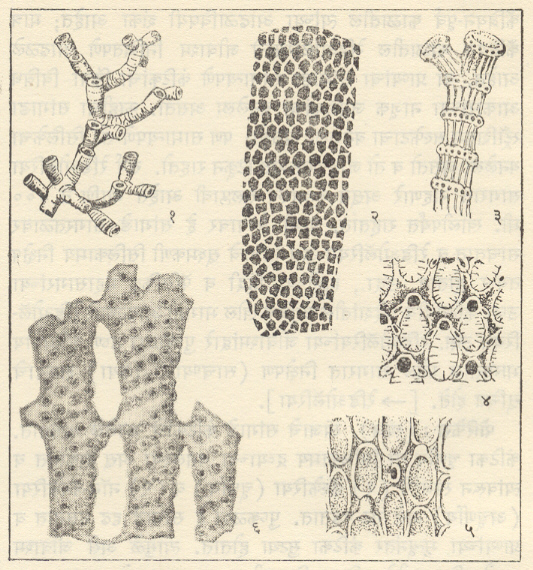
पॉलिझोआ : (ब्रायोझोआ). हे प्राणी मुख्यत्वे सागरात राहणारे व निवहजीवी (समुहांत राहणारे) आहेत. त्यांचे निवह पुढील स्वरूपांत आढळतात : वनस्पतीसारखी वाढ, खडक, दगडगोटे वा इतर प्राण्यांची मुख्यत्वे मॉलस्कांची कवचे यांच्यावरील लेप आणि लहान अथवा मोठे संधितरूपातील पिंड.प्रत्येक प्राण्याची राहण्याची जागा कायलोस्टोमॅटा गटात पेटीसारखी, क्रिप्टोस्टोमॅटा, सायक्लोस्टोमॅटा व ट्रेपोस्टोमॅटा यांच्या गटात नळीसारखी किंवा टेनोस्टोमॅटाप्रमाणे पटलित थरांची बनलेली असते. सामान्यपणे ते चूर्णीय असतात परंतु कायलोस्टोमॅटा आणि टेनोस्टोमॅटा यांच्यातील काही शृंगी (केराटीन या प्रथिनाने युक्त ) असू शकतात. नलिकाकार प्रकारात द्वारके (छिद्रे) टोकाशी असतात. पॉलिझोआ ऑर्डोव्हिसियनमध्ये अवतरले. पर्मियनअखेरीस निर्वंश झालेले ट्रेपोस्टोमॅटा व क्रिप्टोस्टोमॅटा हे पुराजीव काळातील प्रमुख पॉलिझोआ प्राणी होते व ते बहुतकरून भित्ती बनण्याच्या स्थितीशी निगडित होते. सायक्लोस्टोमॅटा पुराजीव काळात थोडेच होते. मात्र मध्यजीव काळात ते जलदपणे विपुल झाले व विशेषतःउत्तर क्रिटेशसमध्ये त्यांचा परमोत्कर्ष झाला. स्टोमॅटोपोरा, इडमोनिया, एंटॅलोपोराहे उत्तर क्रिटेशसमधील महत्त्वाचे वंश होत. कायलोस्टोमॅटा जुरासिकमध्ये अवतरले आणि उत्तर क्रिटेशस व नवजीव काळांत ते अतिविपुल होते. मेंब्रॅनिपोरा. सेलोपोरा, मायक्रोपोरा, क्रिब्रिलिनाहे त्यांचे अधिक महत्त्वाचे वंश होत.
कायलोस्टोमॅटाच्या प्लायोसीन काळात असलेल्या काही जाती हल्ली सागरात राहत आहेत. पॉलिझोआ सामान्यतःउथळ पाण्यात राहणारे आहेत कायलोस्टोमॅटा व टेनोस्टोमॅटांचे काही प्रकार अतिशय खोल ठिकाणीही आढळतात.
अँफिन्युरा व स्कॅफोपोडा (मॉलस्का) : सर्व अँफिन्युरा प्राणी सागरात राहणारे असून ते ऑर्डोव्हिसियन काळापासून पृथ्वीवर असल्याचे दिसून येते.
फक्त पॉलिप्लॅकोफोरा गणाचे ऑर्डोव्हिसियनपासूनचे जीवाश्म आढळतात. हे मुख्यत्वे उथळ पाण्यात राहणारे आहेत, हल्लीच्या काही जाती मात्र खोल पाण्यातही राहतात. या प्राण्यांचे कवच आठ तकटांचे असून तकटे किंचित एकमेकांवर आलेली व अग्र-पश्च दिशेत सरळ मांडलेली असतात. त्यांच्यावर नक्षीकाम असते व ती द्विपार्श्वसममित असतात.हेल्मिंथोकायटान, कार्बॉनिफेरसमधील ग्रिफोकायटॉन व नवजीवातील ⇨ कायटॉन हे अधिक सामान्यपणे माहीत असलेले वंश म्हणता येतील.
स्कॅफोपोडा हा सागरी मॉलस्कांचा दुसरा गट असून ऑर्डोव्हिसियन काळापासूनचे त्याचे जीवाश्म माहीत आहेत मात्र एकूण जीवाश्मांमधील त्याचे स्थान अगदी क्षुल्लक आहे. इओसीनपासून आतापर्यंत आढळणारा डेंटॅलियम हा वंश या गटाचे नमुनेदार उदाहरण आहे. या गटातील प्राण्यांची कवचे शंकूसारखी ते जवळजवळ दंडगोलाकार असतात. हे प्राणी चिखलात वा वाळूमध्ये अंशत पुरलेल्या स्थितीत राहतात व कवचाचे पश्च टोक गाळाच्या वर आलेले दिसते. हे प्राणी किनाऱ्यापासून १,००० मी. वा त्यापेक्षा जास्त खोलीपर्यंतच्या पाण्यात आढळतात.[⟶ मॉलस्का].
ब्रॅकिओपोडा, सिर्रिपिडिया, मॅलॅकोस्ट्रका वगैरे : (आर्थ्रोपोडा). ट्रायलोबिटा व ऑस्ट्रॅकॉडा यांच्या व्यतिरिक्त आर्थ्रोपोडाचे इतर पुष्कळ गट पूर्व पुराजीव काळापासूनच अवतरले होते पण त्यांचे जीवाश्म कमी असून ते चांगले टिकून राहिलेले नाहीत. काही बाबतींतील स्थानिक अपवाद वगळल्यास त्यांच्या स्तरवैज्ञानिक महत्त्वाविषयी अल्पशीच माहिती उपलब्ध आहे म्हणून त्यांच्या मोजदादीपलीकडे इतर कोणती विशेष माहिती देण्याची गरज नाही.
कॅंब्रियन काळापासूनचे ब्रॅकिओपोडा माहीत आहेत परंतु त्यांचे थोडेच जीवाश्म आढळतात. एस्थेरिया ह्या बायव्हाल्व्हियासारखे पृष्ठवर्म असलेल्या वंशाचा केवळ येथे उल्लेख करण्यासारखा आहे. तो वंश डेव्होनियन काळापासून आहे आणि त्यातील प्राणी गोड्या व कधी कधी मचूळ पाण्यात राहतात. [⟶ ब्रॅकिओपोडा].
सिर्रिपिडियाचा हल्लीचा प्रतिनिधी म्हणजे बार्नेकल (कित्येक तुकड्यांनी बनलेल्या चूर्णीय कवचाने संरक्षित असा) बॅलॅनस वंश होय. हे प्राणी सागरात राहाणारे असून बहुतकरून ते किनाऱ्याजवळ समुद्रतटीय पट्ट्यात राहतात. त्यांचे कार्बॉनिफेरस काळापासूनचे जीवाश्म माहीत असून पूर्व क्रिटेशस काळापर्यंत ते वृंताने तळाला चिकटून राहत असत. वृंत नसलेले प्रकार उत्तर क्रिटेशसमध्ये अवतरले व उत्तर नवजीव काळात सर्वसामान्य झाले.
मॅलॅकोस्ट्रॅकाच्या कित्येक गणांपैकी लेप्टोस्ट्रॅका (फायलोकॅरिडा) हा एक गण असून तो सर्वसामान्य कवचधारींचा (क्रस्टेशियनांचा) आदिम गट मानला जातो व त्यातील काही पुराजीव काळातील प्राणी पुष्कळ प्रमाणात सध्याच्या प्राण्यांसारखे होते. हे प्राणी समुद्रात राहणारे आहेत.
आयसोपोडा व अँफिपोडा हे उपगण ट्रायसिकपासून नंतरच्या काळात असल्याचे माहीत असून ते नवजीव काळापासून सर्वसामान्य झाले. ते बहुतेक
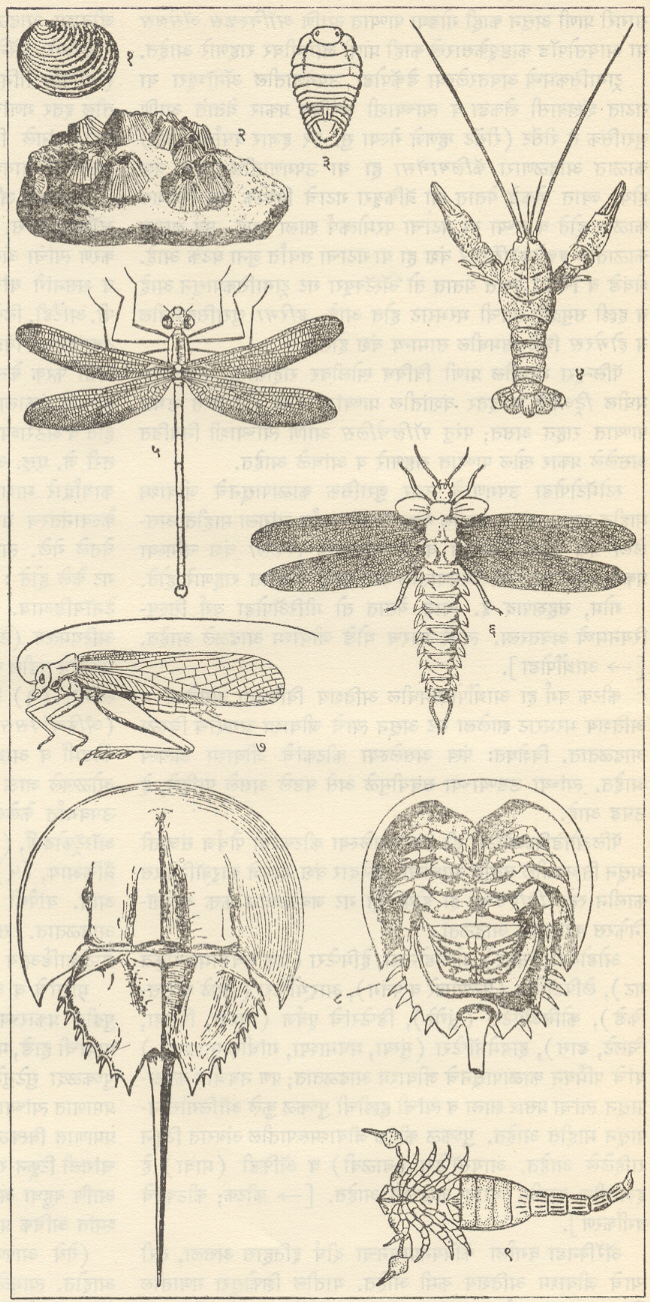
सागरी प्राणी असून काही गोड्या पाण्यात आणि ऑनिस्कस ॲसेलस या आयसोपॉड काष्ठयूकेसारखे काही प्राणी जमिनीवर राहणारे आहेत.
ट्रायसिकमध्ये अवतरलेल्या डेकॅपोडा उपगणातील ॲनॉम्यूरा या गटात शंखवासी खेकडा व त्याच्याशी संबंधित प्रकार येतात आणि जुरासिक ते रीसेंट (रीसेंट म्हणजे गेल्या सु. ११ हजार वर्षांचा काळ) काळात आढळणारा कॅलियानॅसा हा या उपगणातील प्रमुख वंश होय. ज्यात खेकडे येतात त्या ब्रॅकियूरा गटाचे कित्येक वंश क्रिटेशस काळात होते व सध्या या गटाचा परमोत्कर्ष झाला आहे. पूर्व लायस काळातील इओकॉर्सिन वंश हा या गटाचा सर्वांत जुना घटक आहे. शेवंडे व चिमोरे ज्यात येतात तो ॲस्टॅक्यूरा गट ट्रायासिकपासून आहे व हल्ली समुद्रात त्याची भरभराट होत आहे. इरिमा जुरासिकमधील व होमेरस क्रिटेशसमधील सामान्य वंश होत.
पॅलिन्यूरा गटातील प्राणी विविध खोलीवर राहातात. मध्यजीवामधील ट्रिऑन व इतर वंशांतील प्राण्यांना डोळे होते व ते उथळ पाण्यात राहत असत परंतु पॉलिचेलिस आणि त्याच्याशी निगडित असेलेल प्रकार खोल पाण्यात राहणारे व आंधळे आहेत.
स्टोमॅटोपोडा उपगणाचे उत्तर जुरासिक काळापासूनचे जीवाश्म माहीत असून त्या काळातील स्कुल्डा हा सर्वांत चांगला माहीत असलेला वंश होय. क्रिटेशस व इओसीनमध्ये स्क्विला वंश चांगल्या प्रकारे आढळतो. या उपगणातील सर्व प्राणी सागरात राहणारे होते.
गोम, सहस्त्रपाद इ. ज्यात येतात तो मीरिॲपोडा वर्ग सिल्युरियनमध्ये अवतरला. त्याचे फारच थोडे जीवाश्म आढळले आहेत. [ ⟶ आर्थ्रोपोडा].
कीटक वर्ग हा आर्थ्रोपोडामधील अतिशय विविधता असलेला व अतिशय भरभराट झालेला गट असून त्याचे जीवाश्म अगदीच विरळा आढळतात. विशेषत पंख असलेल्या कीटकांचे जीवाश्म अत्यल्प आहेत. त्यांच्या उडण्याच्या सवयींमुळे असे घडले असले पाहिजे, हे उघड आहे.
पॅलिओडिक्टिऑप्टेरा ही पंख असलेल्या कीटकांची पौर्वज संजाती असून तिच्यातील सर्वांत आधीचा नमुनेदार वंश म्हणजे कार्बॉनिफेरस कालीन स्टेनोडिक्टिआ हा होय. हा गट जवळजवळ फक्त कार्बॉनिफेरस काळातच आढळतो.
ओडोनेटा (चतुर), प्रोटोडोनाटा, हेमिप्टेरा (एक सर्वसामान्यीकृत गट), लेपिडॉप्टेरा (फुलपाखरे व पतंग), आर्थोप्टेरा (झुरळे व रातकिडे), कोलिऑप्टेरा (भुंगेरे), डिप्टेरांचे पूर्वज (माश्या, पिसवा, चिलटे, डास), हायमेनॉप्टेरा (मुंग्या, मधमाश्या, गांधील माश्या इ. ) यांचे पर्मियन काळापासूनचे जीवाश्म आढळतात पण नवजीव काळापासून त्यांचा प्रसार झाला व त्यांची हल्लीची पुष्कळ कुले ऑलिगोसीन पासून माहीत आहेत. आयसॉप्टेरा (वाळवी) व ॲफिडी (मावा) हे इओसीन कालीन थरांत आढळले आहेत. [ ⟶ कीटक कीटकांचे वर्गीकरण].
ॲरॅक्निडा वर्गाला कँब्रियनपासूनचा दीर्घ इतिहास असला, तरी त्याचे जीवाश्म अतिशय कमी आहेत. यातील झिफोसूरा गणातील लिम्युलस हा एकमेव वंश सध्या हयात असून किनाऱ्याजवळील समुद्रात यातील प्राणी राहतात. उत्तर जुरासिक काळापर्यंत हा गण संख्येने कमीच होता. यातील पुराजीव कालीन काही प्राणी कदाचित गोड्या पाण्यात राहत होते. ⇨ यूरिप्टेरिडा गणातील प्राणी विंचवासारखे असून इतर आर्थ्रोपोडांच्या मानाने ते अधिक मोठे होते. डेव्होनियन कालीन टेरिगोटस अँग्लिकस हा प्राणी जवळजवळ २ मी. लांब होता व तो ज्ञात असलेला सर्वांत मोठा आर्थ्रोपॉड आहे. स्कॉर्पिओनिडा गण सिल्युरियनमध्ये अवतरला पण त्याचे थोडेच जीवाश्म आढळले आहेत व तेही चांगले टिकून राहिलेले नाहीत. पेडिपॅल्पी, ॲरॅनेइडा, स्यूडोस्कॉर्पिओनिडा (चर्नेटिडिया), फॅलँजिडा (ऑपिलिओनिना), ॲकॅरिना व अँथ्रॅकोमार्टी यांसारख्या ॲरॅक्निडातील इतर गणांचे जीवाश्म आढळले आहेत परंतु ते अल्प असून तेही चांगले टिकून राहिलेले नाहीत म्हणून त्यांचा खास उल्लेख करण्याची आवश्यकता नाही. [ ⟶ ॲरॅक्निडा].
पृष्ठवंशी जीवाश्म गट: मत्स्य वर्ग : [ ⟶ मत्स्य वर्ग ]. ॲरिस्टॉटल (इ. स. पू. ३८४ – ३२२) यांच्या काळापासून माशांचे वर्गीकरण त्यांचा अंतर्गत व बाह्य कंकाल, तसेच पर म्हणजे पक्ष (फिन) व श्वसनांगे यांचे स्वरूप व स्थान यांना अनुसरून केलेले आढळते. पी. आर्टेडी, लिसेपिडी, जी. एल्. क्यूव्ह्ये, ए.व्हालांस्येन यांच्यासारख्या तदनंतरच्या विद्वानांनी उपास्थियुक्त (कूर्चायुक्त) मत्स्य व अस्थिमत्स्य असा फरक केला होता व या प्रत्येकाचे कित्येक उपगट केले होते. दात व कंकालाच्या अवशेषांच्याही रूपात माशांचे जीवाश्म माहीत होते व अठराव्या शतकात त्यांचे वर्णनही केले गेलेले होते. असे असले, तरी जे. एल्. आगास्सिझ (१८०७-७३) यांनी आपल्या संस्मरणीय कार्याद्वारे माशांच्या जीवाश्मांसह एकूण प्राणिजातीचे पूर्ण सर्वेक्षण केल्यानंतरच माशांचे जीवाश्म वर्गीकरणाच्या पद्धतीमध्ये विचारात घेतले गेले. त्यांनी खवल्यांच्या वैशिष्ट्यांनुसार माशांचे पुढील चार गट केले होते प्लॅकोडिआय, गॅनॉइडिआय, सायक्लॉयडिआय आणि टेनॉयडिआय. जे. म्यूलर यांनी या चारांपैकी शेवटच्या दोन गटांचा अस्थिमत्स्य (टेलिऑस्टिआय) हा एक गट केला व उपास्थिमय मत्स्यांचे पुढील चार उपवर्ग केले. (१) लेप्टोकार्डिआय, (२) सायक्लोस्टोमी, (३) सिलॅकिआय आणि (४) डिप्नोई. लेप्टोकार्डिआय (अँफिऑक्सस) हा एक स्वतंत्र वर्गच मानला जातो. तसेच ऑस्ट्रॅकोडर्मी व आर्थ्रोडायरा हे दोन निर्वंश गट स्वतंत्र उपवर्ग म्हणून ओळखले जाऊ लागले आहेत. त्यानुसार मत्स्य वर्गाची पुढील सात उपवर्गात केलेली विभागणी आजकाल योग्य मानली जाते: (१) ऑस्ट्रॅकोडर्मी, (२) सायक्लोस्टोमी, (३) आर्थ्रोडायरा, (४) इलॅस्मोब्रँकिआय, (५)डिप्नोई,(६) गॅनॉइडिआय व (७) टेलिऑस्टिआय. यांपैकी पहिला आणि तिसरा उपवर्ग फक्त पुराजीव काळात आढळतात. सायक्लोस्टोमींचे जीवाश्म आढळले नाहीत, तसेच लेप्टोकार्डिआय उपवर्गाच्या जीवाश्मांचीही नोंद झालेली नाही.
माशांचे व वस्तुतः सर्व पृष्ठवंशी प्राण्यांचे जीवाश्म हे सांगाड्याच्या पुढील प्रकारच्या घटकांच्या रूपात आढळतात. कवटीची व हातापायाची हाडे, मणके, दात, त्वचीय शल्क वा खवले इत्यादी. हे जीवाश्म पुष्कळदा सुटेसुटे आढळतात व कधीकधी त्यांपैकी काही थोड्याफार प्रमाणात त्यांच्या मूळच्या सांधलेल्या स्थितीत आढळतात. तथापि अल्प प्रमाणात विस्कळीत झालेला सांगाडा असणारे किंवा खवलेयुक्त त्वचा चांगली टिकून राहिलेले काही संपूर्ण प्राण्यांचे नमुनेही आढळले आहेत आणि बहुधा असे नमुने इतर पृष्ठवंशीयांच्या मानाने माशांच्या जीवाश्मांत अधिक प्रमाणात आढळतात.
(येथे आपण प्राण्यांच्या गटांचा विचार जीवाश्मांवरून करीत आहोत. त्यामुळे प्राण्यांच्या मऊ भागांपेक्षा त्यांच्या जीवाश्मरूपात टिकून राहिलेल्या भागांकडे अधिक लक्ष दिलेले आहे. त्यामुळे येथे देण्यात आलेली प्राण्यांची पूर्ण सांगाड्याची चित्रे ही उपलब्ध जीवाश्म व बहुतकरून या निर्वंश प्राण्यांशी निगडित असलेल्या जिवंत प्राण्यांची रचना यांच्या आधाराने तज्ञांनी फेरजुळणी करून काढलेली आहेत).
(१) ऑस्ट्रॅकोडर्मी उपवर्ग यूरोप व अमेरिकेच्यासंयुक्त संस्थानांतील पश्चिमेकडील राज्यांतील उत्तर ऑर्डोव्हिसियन कालीनखडकांत आढळलेले सुटे, गाठी गाठी असणारे खवले हे माशांचे सर्वांत जुने जीवाश्म आहेत. ते बहुधा सेफॅलॅस्पिड ऑस्ट्रॅकोडर्मीचे म्हणता येणे शक्य आहे. इंग्लंड व बाल्टिक समुद्रातील अझेल बेटातील उत्तर सिल्युरियन खडकांमध्ये विषमशल्की (हेटेरोस्ट्रॅचीअन) थीलोडस व लॅनॅर्किया
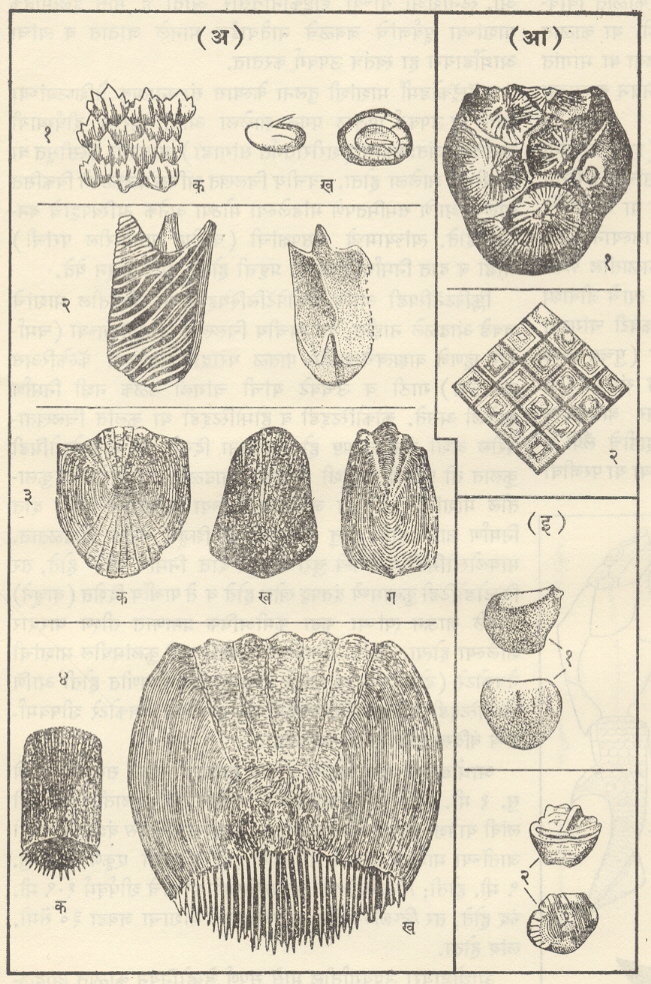
या प्राण्यांची जवळजवळ पूर्ण शरीरे आढळली आहेत. ॲनॅस्पिडा गणातील बिर्किनिया व टेरोलेपीस सारखे काही इतर ऑस्ट्रॅकोडर्मी पॉलिॲस्पिससारखे काही हेटेरोस्ट्रॅसी गणातील (ऑस्टिओस्ट्रॅसू गणातील) सेफॅलॅस्पिस हे जीवाश्म इंग्लंड व उत्तर अमेरिकेच्या सिल्युरियन खडकांत आढळले आहेत परंतु टिकून राहण्यासारख्या कठीण भागांच्या अभावामुळे त्यांच्या नोंदी पूर्ण नाहीत. हल्लीच्या अँफिऑक्सस प्रमाणे त्यांना दीर्घस्थायी पृष्ठरज्जू, अतिशय आदिम प्रकारचा जबडा व केवळ वक्षीय (अंसीय) व श्रोणि-पक्ष (पश्चपक्ष) होते परंतु उपास्थीचे कॅल्सीभवन होण्याच्या (कॅल्शियमाची लवणे साचविण्याच्या)प्रवृत्तीमुळे अपूर्णावस्थेत का होईना प्राणी टिकून राहण्यास काहीशी अनुकूल स्थिती निर्माण होत होती. यावरून फक्त एवढेच दिसून येते की, या सर्वांत जुन्या जीवाश्मांवरून सूचित होणाऱ्या काळाच्या आधीच मासे अवतरले होते.
उत्तर सिल्युरियन व पूर्व डेव्होनियन काळांतील या प्रकारांमध्ये खवले सांधले जाऊन बनलेल्या पट्टांनी डोके व धडाचा वक्षीय भाग यांभोवती चिलखत निर्माण होण्याची प्रवृत्ती आढळते. ॲनॅस्पिडा गणात शरीरावरील खवले नियमित ओळीत मांडलेले असत आणि चिलखत सामान्यपणे लहान बहुभुजाकृती पट्टांचे बनलेले असे. हेटेरोस्ट्रॅसी गणात थीलोडस व लॅनॅर्किया यांसारख्या अधिक आदिम वंशामध्ये शरीरावर गाठीगाठीचे खवले असत परंतु इतरांमध्ये खवले सांधले जाऊन बनलेल्या लहान बहुभुजाकृती पट्टांपासून डोके व धडाचा वक्षीय भाग यांच्याभोवती चिलखत निर्माण होण्याची प्रवृत्ती दिसून येते. ऑस्टिओस्ट्रॅसी गणातील विशेषत सेफॅलॅस्पिस व त्याच्याशी निगडित प्रकारांमध्ये डोके व धडाचा वक्षीय भाग यांच्याभोवतीचे बळकट चिलखत चांगल्या प्रकारे निर्माण झाले होते. अँटिआर्की गणातील प्राण्यांचे चिलखत सममितपणे मांडलेल्या व एकमेकांवर येणाऱ्या मोठ्या पट्टांचे बनलेले होते. या प्राण्यांत वक्षीय भागात पार्श्विक युग्मित उपांगेही निर्माण झाली होती, त्यांच्या शरीरावरील खवले उभ्या दिशेत नियमितपणे ओळीत मांडलेले असत,त्यांच्या कंकालात अस्थिकोश निर्माण झाल्याचेही दिसून येते.
ॲनॅस्पिडा गणातील माशांचे शरीर पोहणाऱ्या माशांप्रमाणे तर्कुरूप होते परंतु इतर ऑस्ट्रॅकोडर्मी माशांची शरीरे चापट व विशेषतः अग्र बाजूस रुंद होती ते मासे बहुतकरून समुद्राच्या तळावर वा तळानजीक राहणारे होते. उपास्थिमय स्परूप आणि जबड्यांचा तसेच वक्षीय वा श्रोणि-पक्षाचा अभाव यांवरून ते जरी मूलतः आदिम असल्याचे सूचित होत असले, तरी डोके व धडाचा वक्षीय भाग यांच्याभोवती कठीण चिलखत निर्माण होणे, हे कठीण संरचनेच्या विकासातील वैशिष्ट्य आहे. या गणातील मासे सर्वसामान्यपणे लहान होते. यांच्यापैकी बरेचसे केवळ काही सेंमी. च लांब होते परंतु त्यांच्यापैकी काही मोठे होते. उदा., पूर्व डेव्होनियन काळातील सेफॅलॅस्पिस मॅग्निफिका या माशाचे शीर्षवर्म ( डोक्याभोवतीचे कायटिनमय कवच) २२ सेंमी. होते व हा तेव्हाच्या विशेष मोठ्या माशांपैकी एक मासा होता.
ऑस्ट्रॅकोडर्मी माशांचे अवशेष बहुतकरून नदीद्वारे व सरोवरात साचलेल्या मृत्तिका, शेल व स्तरभिदुर (थराला अनुसरून भंग पावणारे) चुनखडक यांमध्ये आढळले आहेत. शिवाय यांच्या अवशेषांच्या बरोबर इतर कोणत्याही सागरी अपृष्ठवंशी प्राण्यांचे अवशेष आढळत नाहीत. यावरून या माशांच्या जीवनास नद्यांमधील व जमिनीवरील सरोवरातील गोड्या पाण्यात सुरुवात झाली असल्याचे सूचित होते.
ऑस्ट्रॅकोडर्मी उपवर्गाचे मासे उत्तर ऑर्डोव्हिसियन काळात अवतरले आणि सिल्युरियनच्या अखेरीस व पूर्व डेव्होनियन काळात त्यांचा जलदपणे विकास झाला. जवळजवळ सर्व डेव्होनियन काळात विविधता व विपुलता या दृष्टींनी त्यांची फार वाढ झाली.
या काळात यूरोप, उत्तर अमेरिका, ग्रीनलंड, अंटार्क्टिका व ऑस्ट्रेलिया या भागांत त्यांचा विस्तृत प्रमाणावर प्रसार झाला होता. ते डेव्होनियन काळाच्या शेवटी निर्वंश झाले.
(२) सायक्लोस्टोमी उपवर्ग हल्लीचे लँप्री (इतर माशांना चिकटून त्यांच्यावर जगणारे) व हॅगफिश (इतर माशांच्या शरीरात छिद्र पाडून त्यांची आतडी व मांस यांवर जगणारे) हे या उपवर्गाचे प्रतिनिधी आहेत. त्यांचा कोणताही भाग कठीण नसल्याने त्यांचे जीवाश्म टिकून राहिलेले नाहीत. मध्य डेव्होनियन काळातील पॅलिओस्पॉंडिलस हा सायक्लोस्टोमी उपवर्गातील मानल्यास त्याचे जीवाश्म मात्र आढळले आहेत. पॅलिओस्पॉंडिलस प्रकारात कवटी चांगल्या प्रकारे कॅल्सीभूत झालेली व पृष्ठवंशाला रुधिर-कमान (पुच्छाकडील रक्तवाहिन्या जिच्यात असतात अशी अधर बाजूची संरचना) व तंत्रिका-कमान(मेरुरज्जू जिच्यातून जातो ती अधर बाजूकडील संरचना) तसेच कंटक (काट्यासारखी वाढ) होते. हल्लीचे लँप्री व हॅगफिश हे परजीवी (इतरांवर जगणारे) आहेत. त्यांच्या या परजीवी जीवनप्रणालीत कठीण भागांची गरज नसल्याने जीवनप्रणालीत कठीण भागांची गरज नसल्याने त्यांच्या शरीरात कठीण भाग नसणे हे विशेषीकरण झालेले असू शकेल.
(३) आर्थ्रोडायरा उपवर्ग : या उपवर्गातील माशांचे जबडे डिप्नोई माशांच्या जबड्यांसारखे आहेत. त्यामुळे कोकोस्टिअन मासे हे डिप्नोई गणाचे आहेत, असे जे. एस्. न्यूबेरी (१८२२ – ९२) व ए. एस्. वुडवर्ड (१८६४ – १९४४) यांचे मत होते. तथापि ई. ओ. स्टेनशिओ यांच्या दृष्टिकोनानुसार आता हे मासे इलॅस्मोब्रँक माशांच्या पूर्वजांचे जवळचे नातेवाईक मानले जातात व त्यांचा आर्थ्रोडायरा हा स्वतंत्र उपवर्ग करतात.
ऑस्ट्रॅकोडर्मी माशांशी तुलना केल्यास संरचनात्मक वैशिष्ट्यांच्या दृष्टीने हा उपवर्ग पुष्कळ प्रगत झालेला आहे. पृष्ठरज्जू दीर्घस्थायी असलेला अंतःकंकाल (शरीरांतर्गत सांगाडा) वरकरणी कॅल्सीभूत वा अस्थीभूत झालेला होता. त्वचीय चिलखत अधिक बळकटपणे विकसित झालेले आणि सममितपणे मांडलेल्या मोठ्या अनेक अस्थिपट्टांचे बनलेले होते.त्यांच्यामध्ये अंसपक्षांची (वक्षीय भागावरील परांची) जोडी व दात निर्माण होण्याची प्रवृत्ती होती, असे दिसून येते.
फ्लिक्टिनॅस्पिटी आणि मॅक्रोपेटॅलिक्थिइडी या कुलांतील माशांचे जबडे आढळले नाहीत पण त्वचीय चिलखतावर गॅनॉइनच्या (चर्माद्वारे म्हणजे बाह्यत्वचे खालील पातळ थराद्वारे स्त्रवलेल्या कॅल्केरिअस द्रव्याच्या) गाठी व उंचवटे यांची ठळक नक्षी निर्माण झालेली असते. कोकोस्टिइडी व होमोस्टिइडी या कुलांत चिलखतावरील अशी नक्षी अस्पष्ट होत जाताना दिसते, तर मायलोस्टोमिडी कुलात ती पूर्णपणे नाहीशी झालेली आढळते. कोकोस्टिमिडी कुलातील माशांच्या खालच्या जबड्यात शंकूच्या आकाराचे लहान दात निर्माण झाले होते परंतु पुष्कळदा ते झिजून गेलेले आढळतात. मायलोस्टोमिडी कुलामध्ये चुरा करणारे दात निर्माण झाले होते, तर टिक्टोडोंटिडी कुलामध्ये दंतपट्ट खोल होते व ते पार्श्वीय दिशेत (बाजूने) दाबले जाऊन त्यांच्या कडा कमीअधिक प्रमाणात तीक्ष्ण धारदार झालेल्या होत्या. क्लिक्टिनॅस्पिडी व कोकोस्टिइडी कुलांमधील माशांची नेत्रकोटरे (डोळ्यांच्या खोबणी) शीर्षवर्मातील खोबणीत होती आणि होमोस्टिइडी व मॅक्रोपेटॅलिक्थिइडी या कुलांमध्ये नेत्रकोटरे शीर्षवर्मांमध्ये बंदिस्त होत गेलेली आढळतात.
आर्थ्रोडायरा उपवर्गातील पुष्कळ माशांची लांबी सर्वसामान्यपणे सु.१ मी. होती पण विशेषेकरून कोकोस्टिइडी कुलातील काहींची लांबी यापेक्षा पुष्कळ जास्त होती. उदा., डायनिक्थिस वंशातील काही जातींच्या माशांचे डोके १ मी. लांब होते व त्यांची एकूण लांबी सु. ९ मी. होती टिटॅनिक्थिस आगास्सिझी माशाचे शीर्षवर्म १.९ मी.रुंद होते, तर डिप्लोग्नॅथॉस मायरॅबिलिस माशाचा जबडा ३० सेंमी. लांब होता.
आर्थ्रोडायरा उपवर्गातील मासे संपूर्ण डेव्होनियन काळात आढळतात. परंतु विशेषकरून मध्य व उत्तर डेव्होनियन काळात त्यांच्यात विविधता आली होती, ते विपुल होते व त्यांचा विस्तृत प्रमाणात प्रसार झाला होता. या काळातील त्यांचे जीवाश्म उत्तर अमेरिका, रशिया व इतर यूरोपीय प्रदेशांत आढळले असून ऑस्ट्रेलियातही (न्यू साऊथ वेल्स) ते आढळले आहेत. मायलोस्टोमिडी कुलातील माशांचे जीवाश्म बहुतकरून अमेरिकेच्या संयुक्त संस्थानातील केवळ उत्तर डेव्होनियन काळातील खडकांत आढळतात. आर्थ्रोडायरा उपवर्गातील माशांचे काही सर्वांत चांगले टिकून राहिलेले जीवाश्म जर्मनीतील व्हिल्डुंगन भागातील उत्तर डेव्होनियन कालीन खडकांत सापडले आहेत. डेव्होनियन कालीन माशांपैकी आर्थ्रोडायरा हा एक महत्त्वाचा घटक असून हे मासे विपुलता व विविधता यांसाठी प्रसिद्ध आहेत.
(4) इलॅस्मिब्रँकिआय उपवर्ग : या उपवर्गातील माशांमध्ये पुष्कळ विविधता असून ते सिल्युरिअन ते रीसेंट काळात आढळतात. त्यांचा अंतःकंकाल उपास्थिमय असून जेव्हा त्यांचा बाह्यकंकाल असतो तेव्हा तो बहुतकरून दात व कठीण खवल्यांच्या स्वरूपात आढळतो.युग्मित क्लोम (कल्ले) चांगले वाढलेले व शेपटी समपालिपुच्छी (जिच्यात पृष्ठवंश सरळ शेवटच्या टोकापर्यंत जाऊन पक्ष समसमान विभागला गेलेला असतो अशी) विषमपालिपुच्छी (जिच्यात पृष्ठवंश पुच्छपक्षाच्या वरच्या वा आधीच्या पालीपर्यंत गेलेला असतो अशी), किंवा सेतुपुच्छी (जिच्यात पक्षाचे टोक आखूड झाल्याने द्वितीयक समपालिपुच्छी पुच्छपक्ष निर्माण झालेला असतो अशी) असते. कवटी खंडरहित उपास्थिमय असते व ती कॅल्शियम फॉस्फेटच्या कणांमुळे अंशत कठीण झालेली असते.आधीचे सर्व व मध्यजीव कालीन काही इलॅस्मोब्रँकिआय उपवर्गाच्या माशांना दीर्घस्थायी पृष्ठरज्जू होता, तो उच्च व नंतरच्या प्रकारांमध्ये विविध प्रमाणात कॅल्सीभूत झालेला होता आणि शार्क व राजा माशांमध्ये कशेरुक अक्षाचे पुष्कळदा स्पष्ट खंड पडलेले आढळतात.या उपवर्गातील माशांचे नेहमी आढळणारे जीवाश्मरूप अवशेष म्हणजे दात,मणके व पक्षावरील कंटक हे होत. दात विविध प्रकारचे

असून ते बहुधा चांगले अणकुचीदार व धारदार कडांचे असतात, त्यांना एकच किंवा अधिक दंताग्रे असतात व पुष्कळदा ते चापट अथवा बोथट शंकूच्या आकाराचा माथा असलेल्या पट्टकीय प्रकारचे असतात. दाताची तोंडातील जागा व त्याचे कार्य यांनुसार त्याचा प्रकार बदलतो. ॲकॅंथोडिडी कुलातील काही मासे वगळता इतरांमध्ये दात बंधाद्वारे पक्के झालेले असतात व ते आधार देणाऱ्या जबड्याशी कधीच सांधले गेलेले नसतात. त्यांचे दंतवल्क (दातावरील बाहेरचे लुकण एनॅमल) हे वाहिनी दंतीन (रक्तवाहिन्यांद्वारे झिरपलेले दाताचे द्रव्य व्हॅसोडेंटीन), दंतीन (रासायनिक दृष्ट्या हाडासारखे असणारे कठीण, लवचिक असे दातांचे द्रव्य) किंवा संरचनाहीन दंतवल्काचे म्हणजे गॅनोडेंटीनचे बनलेले असते.
इलॅस्मोब्रँकिआय उपवर्गाची विभागणी पुढील सात गणांत केली जाते: (अ) ॲकँथोडिआय,(आ) प्ल्युरोप्टेरिजिआय, (इ) ऱ्हेनॅनिडी, (ई) स्टेगोसिलॅचिआय, (उ) इक्थिओटोमी, (ऊ) सिलॅचिआय व (ए) हॉलोसेफालाय.
(अ) अँकॅंथोडिआय या गणातील पुष्कळ वंशांची संरचनात्मक तपशीलवार माहिती जवळजवळ पूर्णपणे मिळाली आहे, अगदी थोड्याच वंशांची माहिती सुट्यासुट्या दातांच्या व कंटकांच्या आधारावर रचण्यात आली आहे. अँकॅंथोडिडी कुलातील माशांना एकच पृष्ठीन पक्ष असतो, मात्र या गणातील इतर कुलांतील माशांना दोन पक्ष असतात. या गणातील मासे जे. एल्. आगास्सिझ यांनी आपल्या व्याख्येनुसार मूलतः गॅनॉयडिआय कुलात समाविष्ट केले होते पण आर्. एच्. ट्राक्वाइर, ए. एस्. वुडवर्ड व ओ. जीकेल यांच्यासारख्या तदनंतरच्या शास्त्रज्ञांनी असे दाखवून दिले की, जरी या माशांची त्वचा रूक्षा (शार्कप्रमाणे खरखरीत असलेली त्वचा) नसली, तरी त्यांचा इलॅस्मिब्रँकिआय उपवर्गात स्वतंत्र गण म्हणून समावेश केला पाहिजे.
या गणातील माशांच्या बाह्य कंकालाचे पुष्कळ कॅल्सीभवन झालेले असून त्वचा व कला यांचे फक्त कवटी व वक्षीय भागातच कॅल्सीभवन झालेले आढळते. यांचा पृष्ठरज्जू दीर्घस्थायी असतो. नेत्रकोटरां भोवती दंतिनाच्या पातळ पट्ट्या असतात.धडावरील त्वचीय चिलखत जवळजवळ मांडलेल्या लहान चौकोनी कणांचे बनलेले असून ते पक्षांवर पसरलेले असते. गायरोकँथिडी कुलातील माशांचे शरीर गोलसर व खोलगट असते. डिप्लोकँथिडी, इश्नॅकँथिडी व ॲकॅंथोडिडी या कुलांतील मासे लांबट तर्कुरूप असतात व त्यांना शंकूच्या आकाराचे सूक्ष्म दात असतात. त्रिवेणी-चतुष्क (जंभ-कमानीचा पृष्ठीन भाग ज्या उपास्थीचा बनलेला असतो ती उपास्थी टेरिगोक्वॉड्रेट) व जंभ उपास्थी यांच्या कॅल्सीभूत पट्टाशी दात पक्के सांधले गेलेले असतात किंवा काहींमध्ये दात नसतात. इश्नॅकँथस या वंशात जंभप्रतरसंधीवरील (जोडावरील) दात वलयाकार वा चापाकार असतात. प्रोटोडस वंशातही दात असेच असतात.
या गणाचा काळ उत्तर सिल्युरियन ते पर्मियन असा असून कार्बॉनिफेरस काळात विशेषकरून ॲकँथोडिडी कुलांतील मासे विस्तृत प्रमाणावर आढळतात. प्रोटोडोंटीडी इन्सर्टी सेडिस कुल: आडव्या ओळीत मांडलेले व प्रत्येक ओळीत एकत्र सांधले गेलेले कंटकाग्र, तसेच जवळजवळ घनरूप असे दात प्रोटोडस वंशात असून जंभप्रतरसंधीवरचे दात वलयाकार वा चापाकार असतात. असेच दात इश्नॅकँथिडी कुलांतील इश्नॅकँथस वंशातही असतात व या साम्यामुळे या कुलाचा ॲकँथोडिआय गणात तात्पुरता समावेश केला आहे.
प्रोटोडस व डोलिओडस या वंशाचे जीवाश्म इंग्लंड, अझेल बेट (बाल्टिक समुद्र) व न्यू ब्रन्सविक या प्रदेशांतील डेव्होनियन कालीन खडकांत आढळले आहेत. पॅलिओडस व आर्किडॉन यांचे जीवाश्म पोर्तुगालमधील ऑर्डोव्हिसियन कालीन कोनोडोंटांच्या जोडीने आढळतात. ते बहुतकरून या गणातील असावेत.
(आ) प्ल्युरोप्टेरीजिआय : या गणातील माशांचे दात क्लॅडोडस प्रकारचे असून त्याच्या सपाट तळावर एक प्रमुख दंताग्र व अनेक उपनलिका असतात आणि त्यांच्या कित्येक ओळी एकाच वेळी कार्यान्वित झालेल्या असल्या पाहिजेत. या माशांना दोन बुटके (लहान) पृष्ठीन पक्ष असून त्यांना अग्रीय कंटक नसणे शक्य आहे. वक्षीय व श्रोणिपक्षांना संधियुक्त नसलेल्या समांतर अरीय उपास्थींचा आधार असतो व ते तळाशी अंशतः सांधलेले असतात. कशेरुक-अक्ष बाह्यतः खंडयुत नसतो.
उत्तर अमेरिका व इंग्लंड, आयर्लंड, बेल्जियम यांसारख्या कित्येक यूरोपीय देशांमधील उत्तर डेव्होनियन ते पूर्व पर्मियन काळातील खडकांत या माशांचे जीवाश्म आढळले आहेत. टेनोकँथस कोस्टेलॅटस सारख्या काही थोड्या जातींचा स्कॉटलंडमधील पूर्व कार्बॉनिफेरस कालीन खडकांत जवळजवळ संपूर्ण असा जीवाश्म सापडला आहे. पूर्व कार्बॉनिफेरस काळातील क्लॅडोसिलॅचिआय वंशातील काही जातींतील माशांची लांबी २ मी.पर्यंत होती. या वंशातील व गणातील इतर माशांचे दात, कंटक इत्यादींचे जीवाश्म यूरोपातील पुष्कळ देश, उत्तर अमेरिका व भारतासह काही पौर्वात्य देश या प्रदेशांतील उत्तर डेव्होनियन व पूर्व कार्बॉनिफेरस कालीन खडकांत सापडले आहेत.
(इ) ऱ्हेनॅनिडी : या गणातील माशांची विविधता मर्यादित आहे. यांचे कशेरुक-काय कड्याच्या आकाराचे असतात. कवटीच्या माथ्यावर कमीअधिक प्रमाणात सांधल्या गेलेल्या चर्मपट्टांचे वा गुलिकांचे आच्छादन असते. पश्चकपालास्थि – कंद (पश्चकपालास्थीवरील- डोक्याच्या मागील भागाच्या अस्थीवरील-अस्थीचा पुढे आलेला वाटोळा फुगीर भाग) मोठा व युग्मित असतो. वक्षीय कमानीला थोड्या सडपातळ खंडयुत अरीय उपास्थींचा आधार मिळालेला असतो. या गणातील दोन कुलांपैकी ॲस्टरोस्टीडी कुलाचे जीवाश्म उत्तर अमेरिकेची संयुक्त संस्थाने व प्रशियाच्या ऱ्हाईन नदीलगतच्या प्रदेशातील डेव्होनियन कालीन खडकांत सापडले आहेत. जॅगोरिनिडी कुलाचे जीवाश्म विरळाच आढळतात व ते कवटीचे तुकडे, दात आणि वक्षीय कमान या रूपांत जर्मनीतील उत्तर डेव्होनियन काळातील खडकांतील जीवाश्म गटांत आढळतात.
(ई) स्टेगोसिलॅचिआय : आढळ व विविधता या दृष्टींनी मर्यादित व्याप्ती असलेला हा इलॅस्मोब्रँकिआय उपवर्गाचा आणखी एक गण आहे. या गणाची माहिती आपल्याला बेल्जियम मधील पूर्व कार्बॉनिफेरस कालीन क्रॅस्टोसिलॅचिआय वंशाद्वारे झालेली आहे. त्यांच्या दातांविषयी माहिती उपलब्ध नाही. कवटींच्या थोड्या भागावर सममितपणे मांडलेल्या काही अस्थीभूत पट्टांचे आच्छादन असते. पृष्ठरज्जू दीर्घस्थायी असून अंसपक्षाला तळाशी सांधल्या न गेलेल्या समांतर अरीय उपास्थींचा आधार मिळालेला असतो. काहीसे निश्चितपणे ओळखू येतील असे अवयव असणाऱ्या माशांचा हा या कुळातील एकच वंश असू शकेल.
(उ) इक्थिओटोमी : या गणाचे प्ल्युरॅकँथिडी हे एकमेव कुल आहे. यातील माशांचा अंतःकंकाल पुष्कळ प्रमाणात कॅल्सीभूत झालेला आहे परंतु त्याचे अस्थीभवन झालेले दिसत नाही. नेत्रकोटराभोवती पट्ट नसतात, पृष्ठरज्जू दीर्घस्थायी असतो. तंत्रिका-कमान व रुधिर-कमान लांब व सडपातळ असतात. अंसपक्षाचा अक्ष लांब व खंडयुत असून त्यापासून लहान उपास्थी भिन्न दिशांत गेलेल्या असतात. या माशांचे शरीर सडपातळ असून त्यांच्या दातांना भिन्न दिशांना गेलेली प्रमुख दंताग्रे असतात आणि पृष्ठीन पक्षाला तंत्रिकीय कंटकांपेक्षा (मणक्याच्या काट्यांसारख्या वाढीपेक्षा) संख्येने जास्त असलेल्या सडपातळ कंटकांचा आधार मिळालेला असतो. त्वचा बहुधा रूक्षा नसावी.
प्ल्युरॅकँथस : हा या कुलातील कदाचित सर्वांत जास्त माहिती असलेला वंश आहे आणि यामधील माशांचे जवळजवळ पूर्ण जीवाश्म ऱ्हाईन नदीलगतचा प्रशिया, सायलेशिया व बोहीमिया या भागांत सापडले आहेत. यातील माशांचे डोके अग्र बाजूस अर्धवर्तुळाकार असून त्याच्या पश्चभागी लांब सडपातळ त्वचीय कंटक असतो. इंग्लंड, न्यू साउथ वेल्स, बोहीमिया, सायलेशिया, फ्रान्स, टेक्सस इ. भागातील कार्बॉनिफेरस काळातील दगडी कोळसायुक्त व इतर खडकांत आढळलेल्या विविध वंशातील माशांच्या सुट्या दातांचा समावेश या कुलात केला आहे. हे कुल बहुतकरून ट्रायासिक काळातही अस्तित्वात असावे.
(ऊ) सिलॅचिआय : (शार्क व स्केट). डेव्होनियनमध्ये अवतरलेल्या या कुलातील मासे अजूनही अस्तित्वातात आहेत. या माशांच्या अंतःकंकालाचे कॅल्सीभवन विविध प्रमाणांत झालेले आहे परंतु कलेचे कॅल्सीभवन झालेले नसते. क्लोम-विदर (कल्ल्यातील फटी) उघड्या असतात. अगदी सुरुवातीचे प्रकार वगळता या माशांचा कशेरुक-अक्ष खंडयुत असतो आणि वक्षीय व श्रोणिपक्षांना दोन किंवा तीन आखूड तलोद्भव उपास्थी असतात. या गणात अतिशय विविधता असून त्याचे पुढील दोन उपगण पाडतात. (१) यूसिलॅचिआय : यातील माशांना असंख्य दात असतात व ते जलदपणे येतात आणि पट्टाभ खवले क्वचित सांधले गेलेले असतात. या उपगणाचे दोन गट करतात. (क) क्लोम-विदर पार्श्विक असलेल्या या गटात ११ कुले असून यात शार्क येतात. (ख) क्लोम-विदर अधर बाजूस असलेल्या या गटाची ६ कुले असून स्केट व रे मासे या गटात येतात. (२) ब्रॅडिओडोंटी: या उपगणातील माशांना थोडेच दात असून ते त्यांच्या आयुष्यात सावकाशपणे येतात आणि पट्टाभ खवले सामान्यतः सांधले जाऊन पट्ट बनलेले असतात. यात चार कुले आहेत.
यूसिलॅचिआय उपगणाचा क) गट: एडेस्टिडी, हिबोडोंटिडी व सेस्ट्रॅसिऑनटिडी या कुलांतील माशांचे दात ओरोडस प्रकारचे असतात (आ. २४-३). दाताचा माथा मध्यभागी उंच असतो व अनुदैर्घ्य शिखरापासून अरीय दिशेत गेलेल्या ठळक सुरकुत्या असतात. हिबोडोंटिडी व सेस्ट्रॅसिऑनटिडी कुलांतील माशांच्या दातांच्या माथ्याचा मधला भाग वर येऊन त्याचे प्रमुख दंताग्र बनलेले असते आणि विशेषतः हिबोडसमध्ये व सायनेकोडसच्या काही जातींत खालच्या पातळीवरील अनुदैर्घ्य शिखरांवर दंतिका (लहान दातांसारखे खवले) दिसतात. पृष्ठीन पक्षांना कंटकाचा आधार मिळतो.शेपटी विषमपालिपुच्छी असून पृष्ठरज्जू दीर्घस्थायी असतो.
एडेस्टिडी कुलाचे मासे कार्बॉनिफेरस व पर्मियन काळांतच आढळतात कँपोडस व एडेस्टस हे वंश उत्तर अमेरिका व पुष्कळ यूरोपीय देशांतील पर्मो-कार्बॉनिफेरस (सु. २५ ते २४.५ कोटी वर्षांपूर्वीच्या) काळातील खडकांमध्ये विपुल प्रमाणात आढळतात. हेलिकोप्रियॉन वंशातील माशांच्या प्रतरसंधीच्या भागात दात वलयाकार मांडलेले आढळतात (आ.२४-२). या वंशाचे जीवाश्म उत्तर अमेरिका, रशिया, जपान, कॅनडा, पश्चिम ऑस्ट्रेलिया व भारताच्या कांग्रा भागातील पंजाब-हिमालय या प्रदेशांत विस्तृतपणे आढळतात. हिबोडोंटिडी कुलाचे जीवाश्म डेव्होनियन ते क्रिटेशस या काळातील असून विशेषकरून ट्रायासिक ते क्रिटेशस या काळांत हिबोडस, ॲक्रोडस व ओरोडस हे वंश विस्तृत प्रमाणात आढळले आहेत. सेस्ट्रॅसिऑनटिडी कुल हे ओरोडस प्रकारच्या कंटकाग्र दातांकरिता विशेष प्रसिद्ध आहे. या कुलाचे जीवाश्म जुरासिक ते रीसेंट या काळात आढळले आहेत. चांगली वाढ झालेल्या अशा कंटकाग्र दातांचे चांगले टिकून राहिलेले जीवाश्म (उदा.,सायनेकोडस डयुब्रिसिएन्सिस) इंग्लंडमधील चॉक नावाच्या निक्षेपात आढळले आहेत.
नॉटिडॅनिडी कुलातील माशांच्या दातांना जवळजवळ सपाट अशा बाजूने दाबल्या गेल्यासारख्या तळावर एक वा दोन तीक्ष्ण,अणकुचीदार शंकूच्या आकाराची दंताग्रे असतात. पृष्ठरज्जू बहुधा दीर्घस्थायी व कशेरुक-काय अपूर्ण असतो. पुच्छपक्ष विषमपालिपुच्छी असतो. नॉटिडॅनस हा एक विस्तृतपणे आढळणारा वंश असून तो मध्य जुरासिक ते तृतीय कल्पापर्यंतच्या काळात आढळतो. तृतीय काळात याच्या पुष्कळ जाती आढळल्या आहेत. बव्हेरियामधील उत्तर जुरासिक व लेबाननमधील उत्तर क्रिटेशस काळांतील लिथोग्राफिक खडकांमध्ये या वंशाचे काही उत्कृष्टपणे टिकून राहिलेले जीवाश्म आढळले आहेत. पुष्कळ देशांतील क्रिटेशस व तृतीय कालीन थरांमध्ये या वंशातील माशांचे सुटे दात विस्तृत प्रमाणावर आढळतात.
प्रोटोस्पायनॅसिडी व स्पायनॅसिडी या कुलांतील माशांची शरीरे तर्कुरूप ते काहीशी चापट त्रिफलकीय व मुस्कट रुंद विशालकोनी असते. दात त्रिकोणी किंवा अरुंद भाल्यासारखे दबलेले व वरच्या जबड्यात उभे असे एक दंताग्र असलेले असे असतात. पुच्छपक्ष विषमपालिपुच्छी असतो. प्रोटोस्पायनॅसिडी हे कुल लहान असून त्यातील माशांची रूक्षा दाट असते. या माशाचे जीवाश्म मध्य यूरोपातील जवळजवळ उत्तर जुरासिक कालीन खडकांपुरतेच मर्यादित आहेत. स्पायनॅसिडी कुलाचा काल उत्तर क्रिटेशस ते रीसेंट असा असून त्याचे जीवाश्म मध्यपूर्व, ईजिप्त व बेल्जियम या भागांतील उत्तर क्रिटेशस व इओसीन कालीन खडकांत सापडले आहेत.
प्रिस्टोफोरिडी कुलातील माशांचे धड तर्कुरूप असून मणके संकेंद्रस्तरी (एकच केंद्र असलेल्या वर्तुळाकार उपास्थिमय थरांचे बनलेले) असतात व मुस्कट लांबट असते. सामान्यपणे अणकुचीदार दात असलेली चंचू (चोचीसारखी वाढ) पाहता प्रिस्टोफोरस हा वंश प्रिस्टिडी कुलातील स्क्लेरोऱ्हिंकस वंशासारखा आहे परंतु प्रिस्टोफोरसमध्ये पार्श्वीय उपास्थी नसतात. हा या दोन वंशांतील फरक आहे. प्रिस्टोफोरिडी कुलाचे जीवाश्म लेबाननमधील उत्तर क्रिटेशस व न्यूझीलंडमधील तृतीय कालीन खडकांत आढळले आहेत. या कुलाच्या आधुनिक जाती हल्ली हिंदी महासागरात राहत आहेत. स्क्वाटिनिडी कुलातील माशांना मंक वा एंजल मासे असेही म्हणतात. त्यांचे धड रुंद चापट असते. दात अणकुचीदार शंकूच्या आकाराचे असून त्यांना पार्श्वीय दंतिका नसतात व दंतमूळ खोलगट असते. दातांच्या अनेक ओळी असून त्या सर्व एकाच वेळी कार्य करतात. पंख्याच्या आकाराचे मोठे वक्षीय पक्ष डोक्यावर गेलेले नसतात व पृष्ठीन पक्षांना कंटक नसतात. त्वचेवर लहान पट्टाभ खवले असतात. या कुलाचा काळ जुरासिक ते रीसेंट असा आहे. स्क्वाटिना हा वंश उत्तर जुरासिकपासून आत्तापर्यंत अस्तित्वात आहे. या वंशाचे पूर्णरूपी जीवाश्म बव्हेरियातील लिथोग्राफिक खडकांत व वेस्टफेलियामधील उत्तर क्रिटेशस कालीन खडकांत सापडले आहेत. उत्तर अमेरिकेतील तृतीय काळातील थरांमध्ये या कुलातील माशांचे असंख्य दात व मणके आढळले आहेत.
स्कायलिडी कुलातील मासे तर्कुरूप असून मणके अरस्तरी (बळकट त्रिज्यीय मांडणीच्या पट्टांचे बनलेले) असतात. पृष्ठीन पक्षांना कंटक नसतो व पुच्छपक्ष विषमपालिपुच्छी असतो. दात अनेक, लहान, अणकुचीदार व कित्येक ओळीत असून सर्व एकाच वेळी कार्य करतात. यांचे जवळजवळ पूर्णरूप असे जीवाश्म लेबानन व वेस्टफेलिया येथील उत्तर क्रिटेशस कालीन खडकांत सापडले आहेत. या गटाचे जीवाश्म उत्तर क्रिटेशस आणि तृतीय कालीन खडकांत आढळत असून ते पुष्कळ यूरोपीय देश, उत्तर अमेरिका,उत्तर अफ्रिका, लेबानन या प्रदेशांत सापडले आहेत. भारतामध्ये नर्मदा खोऱ्यातील बाघ थरांमध्ये आढळलेला गिंग्लिमोस्टोमा हा वंश यांचा प्रतिनिधी आहे.
लॅम्निडी कुलातील माशांचे वैशिष्ट्य म्हणजे बळकट कंटकाग्र दात होय. काही वंशात दंतिका असतात व कॅरकॅरोडॉन सारख्या काही वंशांत दातांच्या कडा दंतूर असतात. ओडोंटॅस्पिस वंशात दंताग्र सडपातळ, लांब असून मूळ खोल द्विभिन्न (दोन भाग झालेले) असते. पुष्कळ देशांतील उत्तर क्रिटेशस व पूर्व तृतीय कालीन खडकांमध्ये स्कॅपॅनोऱ्हिंकस, ओडोंटॅस्पिस, ओटोडस, फोरॅक्स, ऑक्सिऱ्हिना, लॅम्ना व कॅरकॅरोडॉन या वंशांचे जीवाश्म विशेषकरून विस्तृतपणे आढळतात आणि ते स्तरवैज्ञानिक सहसंबंध निश्चित करण्यासाठी व जमिनीची पुराभौगोलिक जोडणी करण्याच्या दृष्टीने उपयुक्त ठरले आहेत. कॅरकॅरोडॉन हा मायोसीन व प्लायोसीन काळातील अतिशय महत्त्वाचा वंश आहे. भारतामध्ये नर्मदा खोऱ्यातील बाघ थरांमध्ये व दक्षिण भारतातील उत्तर क्रिटेशस कालीन खडकांत वरील वंशांचे जीवाश्म आढळले आहेत. पंजाबातील प्लायोसीन काळातील शिवालिक शैलसमूहात ओडोंटॅस्पिस वंशाचे जीवाश्म आढळले आहेत. राजस्थानातील बारमेर जिल्ह्यातील कापुर्डीच्या इओसीन कालीन मुलतानी मातीत व बारिपाडाच्या मायोसीन कालीन निक्षेपांत ओडोंटॅस्पिस, कॅरकॅरोडॉन व ऑक्सिऱ्हिना या वंशांचे जीवाश्म सापडले आहेत.
कॅरकॅरीइडी कुलातील माशांचे दात त्रिकोणी,अणकुचीदार,पोकळ दंताग्रे आणि सरळ वा दंतुर कडा व सूक्ष्म दंतिका असलेले असतात. या माशांची दंताग्रे पुष्कळदा किंचित तिरपी असतात लॅम्निडी कुलात तशी नसतात. कशेरुक-काय अधिवृत्त (एका केंद्राभोवती कॅल्शियमी द्रव्याची कित्येक कडी असलेल्या) प्रकारचे असतात व कमानीच्या तळाशी कॅल्सीभूत न झालेल्या उपास्थीच्या पाचरी असतात. पुच्छपक्ष विषमपालिपुच्छी असतो. पृष्ठीन पक्षांना कंटक नसतो. या कुलाचा सर्वसामान्य काळ तृतीय ते रीसेंट असा आहे परंतु दक्षिण भारत व नर्मदा खोऱ्यातील बाघ थर यांमधील उत्तर क्रिटेशस कालीन खडकांमध्ये कॅरकॅरीअँस व प्रॉयॉनोडॉन या वंशांचे जीवाश्म सापडले आहेत. कोरॅक्स व आयस्यूरस हे वंशही दक्षिण भारतातील उत्तर क्रिटेशस कालीन खडकांमध्ये आढळले असून कॅरकॅरीओलॅम्ना, आयस्यूरस, अप्रॉयॉनोडॉन, हायपोप्रायॉन, प्रायॉनोडॉन व स्कोलिओडॉन हे वंश ओरिसातील बालासोर भागातील मायोसीन कालीन खडकांत आढळले आहेत. कॅरकॅरीओलॅम्ना व गॅलिओसेर्डा हे वंश कच्छातील मायोसीन कालीन खडकांत आढळले आहेत. कॅरकॅरिॲस व त्याच्याशी निगडित असलेले जीवाश्म तृतीय कालीन खडकांमध्ये विस्तृतपणे आढळले आहेत.
यूसिलॅचिआय उपगणाचा (ख) गट: स्केट व रे मासे या गटात येतात. त्यांचे क्लोम-विदर अधर बाजूला असतात व सामान्यतः पट्टाभ खवले सांधले जाऊन पट्ट बनलेले असतात. टिकोडोंटिडी व मायलिओबॅटिडी कुलांत दाताचा माथा कंटकयुक्त व चौरसाकृती असतो व ते पट्टकीय रूपात मांडलेले असतात. राजिडी व ट्रायगोनिइडी कुलांत दात लहान,द्विभिन्न मूळ व समभुज चौकोनी किंवा बहुभुजाकृती माथा असलेले असतात व त्यांची जबड्यांतील मांडणी पट्टकीय असते. प्रिस्टिडी व ऱ्हिनोबॅटिडी कुलांमध्ये दात सूक्ष्म, कमीअधिक प्रमाणात बोथट असून एकाच वेळी कार्य करणाऱ्या त्यांच्या कित्येक ओळी असतात. कशेरुक अधिवृत्त प्रकारचे असतात. ऱ्हिनोबॅटिडी व प्रिस्टिडी कुलांतील माशांचे धड दबलेले व लांबट असते, तर इतर कुलांतील माशांचे धड कमीअधिक प्रमाणात बिंबाभ (चकतीसारखे) असते. अंसपक्ष मोठे व डोक्यास जोडले गेलेले असून अग्र बाजूस पसरलेले असतात टॉर्पिडनिडी कुलामध्ये तर अंसपक्ष मुस्कटाच्या अग्र बाजूपर्यंत पसरलेले असतात. व राजिडी कुलात ते श्रोणिपक्षापर्यंत मागे गेलेले असतात. प्रिस्टिडी कुलांतील माशांचे मुस्कट काहीसे चोचीप्रमाणे व तलवारीप्रमाणे असून प्रत्येक बाजूला त्वचीय काटेरी दात असतात पण ते खोबणीत नसतात.
टिकोडोंटिडी हे कुल क्रिटेशस काळातच अस्तित्वात होते व त्या काळातील माशांमधील हे महत्त्वाचे मासे आहेत. यांपैकी टिकोडस वंश यूरोप, उत्तर अमेरिका, जपान, सिरिया, भारत इ. भागांत विस्तृतपणे आढळतो. ऱ्हिनोबॅटिडी कुल उत्तर जुरासिकमध्ये अवतरले आणि इतर कुले उत्तर क्रिटेशसमध्ये अवतरली व अजूनही अस्तित्वात आहेत परंतु ही कुले पुष्कळ देशांत उत्तर क्रिटेशसमध्येच अधिक विपुल प्रमाणात होती. भारतामध्ये टिकोडस वंश नर्मदा खोऱ्यातील बाघ थरांत व दक्षिण भारतातील क्रिटेशस कालीन खडकांत आढळतो. ऑंकोप्रिस्टिस वंश बाघ थरांत आढळला असून मायलिओबॅटिस वंशाचे जीवाश्म कच्छातील इओसीन आणि बालासोर व बारिपाडा(ओरिसा) या भागांतील मायोसीन कालीन खडकांमध्ये सापडले आहेत.
उपगण ब्रॅडिओडोंटी: या गटात अनेक वंश असून मुख्यत्वे सुट्या दातांच्या अथवा दंतपट्टाला चिकटलेल्या दातांच्या गटांच्या रूपात आढळलेल्या जीवाश्मांवरून हे वंश ठरविण्यात आले आहेत. दलन करणारे व चुरडणारे असे या दातांचे प्रकार असून प्रत्येक जंभावर त्यांच्या दोन अथवा अधिक ओळी आढळतात. ते आडव्या दिशेत लांबट असून अग्र-पश्च दिशेत दबले गेलेले असतात. त्यांचे माथे कार्बॉनिफेरस कालीन सॅमोडोंटिडी आणि डेव्होनियन-कार्बॉनिफेरस काळातील कोपोडोंटिडी कुलातल्याप्रमाणे सूक्ष्म सुरकुत्या किंवा छिद्रे असलेले असू शकतात अथवा उत्तर डेव्होनियन ते पर्मियन कालीन कॉक्लिओडोंटिडी कुलातल्याप्रमाणे दाताच्या माथ्यावर उंचवटे असू शकतात. या उपगणातील काही माशांच्या डोक्यावर व धडावर थोडेफार वक्र असे कंटक होते. सॅमोडोंटिडी कुलातील बहुतेक वंशांमध्ये दातांच्या एकाच वेळी कार्य करणाऱ्या कित्येक ओळी होत्या परंतु या ओळी कमी होण्याची प्रवृत्ती आढळते आणि पेटॅलोऱ्हिंकस व जॅनासा यांसारख्या काही विशेषीकृत वंशांमध्ये तर एका वेळी दातांची एकच ओळ कार्य करीत असे. हा उपगण फक्त उत्तर पुराजीव कालीन खडकांतच आढळतो. उत्तर अमेरिका आणि पुष्कळ यूरोपीय व पौर्वात्य देशांत या उपगणाचे जीवाश्म आढळले आहेत. भारतामध्ये कांग्रा भागातील पूर्व कार्बॉनिफेरस काळातील खडकांत कॉक्लिओडोंटिडी कुलातील हॉलोडस वंशाचे जीवाश्म सापडले आहेत.
(ए) हॉलोसेफालाय : पुढील दोन वैशिष्ट्यांमुळे इलॅस्मोब्रँकिआयमधील हा एकमेवाद्वितीय गण आहे: (१) कमानीपेक्षा पुष्कळ जास्त उपास्थिमय कडी असलेल्या कशेरुक-अक्षाचे खंडीभवन अपूर्ण असणे व (२) वरचा जबडा कवटीशी सांधला गेलेला असणे. या माशांचे धड शार्कप्रमाणे असते, मात्र दातांची मांडणी अतिशय विलक्षण असते. खालच्या जबड्यात जंभाच्या प्रत्येक बाजूस एक व वरच्या जबड्यात एक ते तीन मोठे दंतपट्ट असतात. पृष्ठरज्जूच्या भोवती काही प्रमाणात कॅल्सीभवन झालेली कित्येक उपास्थिमय कडी असतात. अंसपक्ष व श्रोणिपक्षांना तळाशी दोन किंवा तीन उपास्थी असतात. स्क्वॅलोरेइडी हे कुल फक्त पूर्व जुरासिकमध्ये आढळते. यातील माशांचे धड लांबट व मुस्कट लांब सडपातळ होते. त्यांना पृष्ठपक्ष नव्हते व अनेक शंक्वाकार वा ताराकृती त्वचीय गाठी होत्या. इंग्लंडबाहेर या कुलाविषयी थोडीच माहिती मिळाली आहे. मायरियाकँथिडी कुलाचे थोडेच जीवाश्म इंग्लंड, बव्हेरिया, जर्मनी इ.प्रदेशांतील जुरासिक कालीन खडकांत सापडले आहेत. या माशांचे धड लांबट होते व डोक्यावर थोडे चर्मपट्ट होते. दंतपट्टाशिवाय त्यांना जंभाच्या प्रतरसंधीवर बळकट कृंतक दात होते. कीमेरिडी हे कुल जुरासिकपासून रीसेंटपर्यंतच्या काळात आढळत असून हल्लीही सागरात या कुलातील मासे आहेत. त्यांचे धड शार्कप्रमाणे लांबट असून खालच्या जबड्यातील प्रतरसंधीवरील दंतपट्टांची एक जोडी चोचीप्रमाणे असते. अंसपक्ष अतिशय मोठे असून त्यांना कंटक असतात व ते उपास्थिमय तळाशी सांधले गेलेले असतात. त्वचा अनावृत्त (खवले वगैरे नसलेली) असते किंवा तिच्यावर रूक्षेच्या कणिका असतात. या कुलाचे जीवाश्म पुष्कळ यूरोपीय देश, न्यूझीलंड, ऑस्ट्रेलिया व अमेरिकेची संयुक्त संस्थाने या भागांतील मध्यजीव व तृतीय कालीन खडकांत सापडले आहेत.
(५) डिप्नोई उपवर्ग : (फुप्फुसमीन). या उपवर्गातील मासे व हल्लीचे इतर मासे यांत एक फरक आहे तो म्हणजे या माशांना फुफ्फुसाचे काम करणारा एक वाताशय (आतड्यापासून अंधनालाच्या स्वरूपात उत्पन्न झालेली वायूने भरलेली पिशवी)असतो व त्यामुळे ते बराच काळ पाण्याबाहेर राहू शकतात. हे वैशिष्ट्य तसेच त्यांच्या हृदयाची व अंतर्गत नासाविवरांची संरचना यांच्यामुळे हे माशांसारखे उभयचर (वा सायरन) मानले जात होते परंतु ऑस्ट्रेलियातील नद्यांमध्ये सेरॅटोडस फॉर्स्टेरी ही जाती आढळल्यामुळे टी.एच्.हक्स्ली यांनी पूर्वी सुचविल्याप्रमाणे त्यांचा क्रॉसेप्टेरिजीआयच्या पूर्वजांशी निकटचा संबंध असावा, हे म्हणणे सिद्ध झाले. शिवाय त्यांच्या जबड्यांची स्वनिलंबित (कवटीशी सरळ सांधल्यामुळे जंभकमानींना स्वतःचा आधार असतो अशी) मांडणी व वैशिष्ट्यपूर्ण दंतविन्यास (दातांची मांडणी) यांच्यामुळे ते इतर माशांपासून वेगळे आहेत असे मान्य झाले आणि त्यामुळे आता त्यांचा स्वतंत्र उपवर्ग केला जातो.
या माशांतील कशेरुक कमानी, फासळ्या व पक्ष यांचे आधार वरवर अस्थीभूत झालेले असतात व अशा तऱ्हेने त्यांना असंख्य कलाजात अस्थी (उपास्थीच्या टप्प्यातून न जाता सरळ कलेमध्ये निर्माण झालेली हाडे) असतात परंतु असे असले, तरी त्यांचा अंतःकंकाल मुख्यत्वे उपास्थिमय असतो. डिप्टेरिडी कुलात कवटीचा माथा कॉस्मीनचे (कोशिकारहित दंतिनासारख्या द्रव्याचे) आवरण असलेल्या हाडांचा बनलेला असतो परंतु इतर कुलांमध्ये हाडांवर लुकण (एनॅमल) नसते. त्यांच्या दोन्ही जबड्यांत दंतपट्ट असून त्यांचे चुरडणारे दात बळकट होते. फॅनेरोप्ल्युरिडी कुलातील माशांना जंभिका व अग्रजंभिकांवर (वरच्या जबड्यातील भागांवर) दातांची ओळ होती. सेरॅटोडोंटिडी आणि टेनोडोंटिडी कुलांत सीमावर्ती दात नव्हते परंतु सेरॅटोडोंटिडीमध्ये हलास्थीवर (नाकाच्या भागातील हाडावर) कापणाऱ्या दातांची जोडी होती. यूरोनेमिडी कुलात लहान शंक्वाकार बोथट दात होते, तर लेपिडोसायरनिडी कुलातील माशांच्या दंतपट्टांना स्वतंत्र कटक (उंचवटे) होते.
खवले चक्रज (ज्यांच्या मोकळ्या कडा सारख्या वाकलेल्या असतात असे) असून ते अंशत एकमेकांस झाकणारे असतात व उघड्या पडलेल्या भागावर कॉस्मीनचे आवरण असते. फेनेरोप्ल्युरिडी कुलात खवल्यांचे बारीक कंटकांत रूपांतर झालेले असते आणि टेनोडोंटिडी कुलांत त्यांच्यावर जालकांचे आवरण असते.
अंसपक्ष व श्रोणि-पक्ष वल्ह्याच्या आकाराचे असून त्यांचा अक्ष लांब, खंडयुत उपास्थिमय असतो आणि लेपिडोसायरनिडी कुलात हे पक्ष दंडाकार झालेले आढळतात. मध्यपक्ष पुच्छपक्षाशी थोडाफार सलग असतो व तो यूरोनेमिडी, टेनोडोंटिडी व सेरॅटोडोंटिडी कुलांमध्ये समपालिपुच्छी किंवा सेतुपुच्छी आणि फॅनेरोप्ल्युरिडी कुलात विषमपालिपुच्छीही असतो पण डिप्टेरिडी कुलात पुच्छपक्ष विषमपालिपुच्छी आणि पृष्ठीन पक्ष व गुदपक्ष स्वतंत्र असतात.
डिप्नोई उपवर्गाचा काळ डेव्होनियन ते रीसेंट असा असला, तरी हा उपवर्ग मुख्यत्वे पुराजीव व मध्यजीव काळांतील आहे. डिप्टेरिडी व फॅनेरोप्ल्युरिडी ही कुले केवळ डेव्होनियनमध्ये आढळतात. टेनोडोंटिडी आणि यूरोनेमिडी या कुलांची भरभराट कार्बॉनिफेरस व पर्मियन काळांत झाली व कदाचित ती ट्रायासिकमध्येही होती. सेरॅटोडोंटिडी कुलाचा काळ ट्रायासिकपासून आत्तापर्यंत असा असून त्याचा मध्यजीव काळात यूरोप, उत्तर अमेरिका, दक्षिण आफ्रिका, भारत, ऑस्ट्रेलिया इ. भागांत विस्तृत प्रमाणावर विस्तार झाला होता. भारतामध्ये या कुलाचा सेरॅटोडस हा वंश उत्तर गोंडवनी गटातील मालेरी, टिकी व येर्रापल्ली थरांत आढळतो, तृतीय काळापासून आत्तापर्यंत या कुलाचा बराच ऱ्हास झाला. त्यांपैकी बहुतकरून सेरॅटोडस हा एकच वंश आत्ता अस्तित्वात असून तोही ऑस्ट्रेलियातच आढळतो. लेपिडोसायरनिडी हे एक काहीसे मर्यादित विस्तार असलेले कुल आहे व त्याचे थोडेसेच जीवाश्म उत्तर व पूर्व आफ्रिकेतील तृतीय कालीन खडकांत सापडले आहेत. हल्ली या कुलातील मासे दक्षिण अमेरिका व आफ्रिकेच्या उष्ण कटिबंधीय भागांतील गोड्या पाण्यात आढळतात. दक्षिण अमेरिकेत लेपिडोसायरन तर आफ्रिकेत प्रोटोप्टेरस वंश आढळतात. [⟶ डिप्नोई].
(६) गॅनॉइडिआय उपवर्ग : या माशांच्या शरीरावरील चर्म आच्छादन हे वैशिष्ट्यपूर्ण अशा गॅनॉइड म्हणजे आकाचाभ (अस्थिमय जाड थरावर एनॅमलचा थर असलेल्या) खवल्यांचे बनलेले असते. हे खवले उखळीच्या सांध्याने जोडले गेलेले असतात. काहींचे खवले मात्र चक्राभ व कंकताभ (मोकळी कडा फणीसारखी दातेरी असते अशा) खवल्यांप्रमाणे एकमेकांवर आलेले आढळतात. कॉंड्रॉस्टेऑयमध्ये पृष्ठवंश उपास्थिमय असतो परंतु क्रॉसोप्टेरिजीआय व इतरांमध्ये कमानी, कटक इत्यादींचे अस्थीभवन वाढत्या प्रमाणात झालेले आढळते आणि क्रॉसोप्टेरिजीआयमधील पॉलिप्टेरिडी व प्रोटोस्पॉंडायलीतील लेपिडोस्टेइडी कुलात अस्थीभवन पूर्ण झालेले असते. कवटीचेही वाढत्या प्रमाणात अस्थीभवन झालेले आढळते. क्रॉसोप्टेरिजीआयमध्ये व पुराजीव कालीन कॉंड्रॉस्टिआय व प्रोटोस्पॉंडायलीमध्ये टेलिऑस्टिआयमधल्याप्रमाणे (अस्थिमत्स्यांच्या उपवर्गातल्याप्रमाणे) अस्थीभवन झालेले असते. गॅनॉइडिआय उपवर्गातील विविध गणांतील वक्षीय कमानीच्या संरचनेवरून डिप्नोई, इलॅस्मोब्रॅंकिआय व टेलिऑस्टिआय यांच्यातील मध्यस्थ दुवा सूचित होतो. अंसफलक (खांद्याचे हाड), अंसतुंड (अंसफलक व उरोस्थी-छातीचे हाड-यांच्यमधील हाड) इत्यादींशी तुल्य अशा लहान संरचना कॉंड्रॉस्टिआय व बहुधा क्रॉसोप्टेरिजीआयमधील जीवाश्मरूप माशांमध्ये उपास्थिमय असतात परंतु इतरांत त्या अस्थिमत्स्यांसारख्या असतात.
या साम्यांमुळे रिचर्ड ओएन (१८०४–९२), ई. डी. कोप (१८४०-९७) व ए. एस्. वुडवर्ड (१८६४–१९४४) यांसारख्या शास्त्रज्ञांनी गॅनॉइड मासे अस्थिमत्स्य यांचा टेलिऑस्टोमी नावाखाली एकत्र समावेश केला आहे. परंतु एकूणच त्वचीय व अंतःकंकालाच्या वैशिष्ट्यांतील या बदलांच्या बरोबर अंतर्गत संरचनांच्या वैशिष्ट्यांतील फरक लक्षात घेतल्यावर गॅनॉइड माशांचा अस्थिमत्स्यांपेक्षा स्वतंत्र उपवर्ग केला जातो.
कवटीच्या हाडांची मांडणी,जंभांची गुंतागुंतीची संरचना, दातांच्या दंतिनस्तराचे आत दुमडले जाणे व क्रॉसोप्टेरिजीआयमध्ये आढळतात त्याचप्रमाणे हलास्थीवरील दातांची वाढ इ.गोष्टी अगदी स्टेगोसेफॅलिया उभयचरांतही असतात. ऑस्टिओलेपिडीतील शिश्नरंध्र व क्रॉसोप्टेरिजीआयतील पॉलिप्टेरिडी कुलातील माशांच्या डोळ्यांभोवतीच्या श्वेत पटलांचे कडे हेही स्टेगोसेफॅलियात आढळते. अशा तऱ्हेने सर्व माशांपैकी क्रॉसोप्टेरिजीआय माशांचे आधीच्या स्टेगोसेफॅलियांशी सर्वात जवळचे संबंध आहेत. व हे दोन्ही प्राणीगट समकालीन होते. तेंव्हा बहुधा क्रॉसोप्टेरिजीआय माशांपासून स्टेगोसेफॅलियांचा पौर्वज गट आला असावा व त्यापासून पुराजीव काळात स्टेगोसेफॅलिया अवतरले असावेत.
हल्लीचे गॅनॉइड मासे बहुतकरून गोड्या पाण्यात राहतात. जरी डेव्होनियन व तृतीय काळांतील सापडलेले गॅनॉइड मासे गोड्या पाण्यात साचलेल्या निक्षेपांतील असले, तरी गॅनॉइड माशांचे बहुसंख्य जीवाश्म सागरी निक्षेपांतील आहेत. उत्तर पुराजीव, ट्रायासिक व जुरासिक काळांमध्ये गॅनॉइड माशांचा कमाल विकास झाला.क्रिटेशस काळापासून अस्थिमत्स्यांनी त्यांच्यावर अखंडपणे मात केलेली आढळते.
गॅनॉइडीआयच्या चार गणांपैकी हेलिकोस्टोमी गण फक्त मध्यजीव काळांत आढळतो. क्रॉसोप्टेरिजीआय माशांचा प्रसार मध्यजीवापेक्षा पुराजीव काळात अधिक प्रमाणात झाला होता, मात्र त्यांपैकी सीलॅकँथिडी कुलाचा प्रसार मध्यजीव काळात विशेष विस्तृतपणे झालेला होता. भारतामध्ये प्राणहिता-गोदावरी खोऱ्यातील जुरासिक कालीन कोटा थरांमध्ये (उत्तर गोंडवनी संघ) इंडोसीलॅकँथस वंशाचे जीवाशम सापडले आहेत [⟶ लॅटिमेरिया]. कॉंड्रॉस्टेऑय माशांमध्ये डेव्होनियन ते जुरासिक काळात ठळकपणे विविधता आली व त्यांचा विस्तार झाला. तदनंतर त्यांचा ऱ्हास होत गेलेला आढळतो. भारतामध्ये त्यांच्यापैकी सॉरिक्थिस, अँब्लिप्टेरस हे जीवाश्म काश्मिरातील पूर्व गोंडवनी संघाच्या पूर्व पर्मियन कालीन गँगॅमोप्टेरीस थरांमध्ये व कदाचित बिहारातील दगडी कोळशाच्य क्षेत्रातील पूर्व ट्रायासिक कालीन पांचेट थरांतही आढळतात. गॅनॉइडी उपवर्गापैकी प्रोटोस्पॉंडायली व त्यातीलही विशेषतः सेमिऑनोटिडी व पिक्नोडोंटीडी कुलांचा प्रसार व विविधता यांच्या दृष्टीने प्रथम क्रमांक आहे. भारतामध्ये सेमिऑनोटिडी कुलाचे लेपिडोटस, डॅपेडियस व टेट्रागोनोलेपिस वंश प्राणहिता-गोदावरी खोऱ्यातील पूर्व जुरासिक कालीन कोटा थरांमध्ये आणि स्फीरोडस वंश त्रिचनापल्लीच्या उत्तर क्रिटेशस कालीन थरांत आढळतो. पिक्नोडोंटीडी कुलातील पिक्नोडस वंशाचे जीवाश्म मध्य प्रदेशातील उत्तर क्रिटेशस कालीन लॅमेटा थरांत व त्रिचनापल्लीच्या उत्तर क्रिटेशस कालीन थरांमध्ये आढळतात. सीलोडस वंशाचे जीवाश्म प्राणहिता-गोदावरी खोऱ्यातील पूर्व जुरासिक कालीन कोटा थरांत व नर्मदा खोऱ्यातील क्रिटेशस कालीन बाघ थरांत व आसामातील गारो टेकड्यांतील इओसीन कालीन सिजू थरांत सापडतात. लेपिडोस्टेइडी कुलातील लेपिडोस्टियस वंशाचे जीवाश्म मध्य प्रदेशातील उत्तर क्रिटेशस कालीन लॅमेटा थरांत आणि मध्य प्रदेशातील राजमहेंद्री जवळील कोटेरू येथे उत्तर क्रिटेशस (वा पूर्व इओसीन) काळातील अंतरा-ट्रॅपी थरांमध्ये आढळतात. तसेच बेलॅनोस्टोमस वंशाचे जीवाश्म मध्य प्रदेशातील लॅमेटा थरांत आढळले आहेत.
(७) टेलिऑस्टिआय उपवर्ग : उत्तर जुरासिक या उपवर्गातील मासे म्हणजे अस्थिमत्स्य अवतरले, क्रिटेशसमध्ये ते चांगले प्रस्थापित झाले, उत्तर क्रिटेशसमध्ये त्यांनी गॅनॉइड माशांना मागे टाकले तेव्हापासून हा सर्व माशांमधील प्रमुख गट झाला आहे. सर्व जलचर पृष्ठवंशी प्राण्यांत अस्थिमत्स्यांनी गोड्या पाण्यात आणि सागरी परिस्थितीत यशस्वीपणे सर्व प्रकारच्या परिस्थितींशी सर्वांत जास्त जुळवून घेतले आहे. त्यांचा विकास व प्रसार विविध दिशांनी होत गेला व त्यांचे पुष्कळ विविधतापूर्ण प्रकार व वंशशाखा निर्माण झाल्या.
त्यांचा अंतःकंकाल पूर्णतया अस्थीभूत किंवा हाडाचा असतो. जंभ साधा असून प्रत्येक शाखा दोन किंवा तीन घटकांची बनलेली असते. गुद व पृष्ठीन पक्षाच्या आधारांची संख्या त्वचा-अरांइतकी (पक्षाला आधार देणाऱ्या अस्थिमय कंटकांइतकी) असते. त्यांच्या त्वचेवर पातळ,लवचिक,चक्राभ वा कंकताभ खवले व क्वचित अस्थिपट्ट असतात. पुच्छपक्ष सामान्यतः समपालिपुच्छी असतो व खवल्याचे टेकू नसतात.
यांचे जीवाश्म सुस्थितीत टिकून राहिलेले असतात व ते शरीराच्या तुकड्यातुकड्यांच्या रूपात आढळतात. यामुळे त्यांचे पुराजीववैज्ञानिक दृष्टीने वर्गीकरण जरा स्थूलमानानेच केलेले असते. त्यामुळे १९४०च्या सुमारास टेलिऑस्टिआय उपवर्गाचे ९ गण व त्यांची ६८ कुले केलेली होती. पुष्कळ जीवाश्मांचे हल्ली अस्तित्वात असलेल्या कुलांतील माशांशी जवळचे संबंध आहेत, असे स्पष्ट दिसते व त्यामुळे निर्वंश झालेल्या जाती वगळता नवजीव काळातील पुष्कळ माशांचे हल्लीच्या कुलांच्या संदर्भातील वर्गीकरणामधील पद्धतशीर स्थान ठरविणे शक्य आहे. अलीकडील संशोधन कार्यामुळे अस्थिमत्स्यांच्या जीवाश्मांबद्दलच्या त्यांच्या वर्गीकरणातील स्थानाच्या संदर्भातील माहितीत पुष्कळ भर पडली असली, तरी त्याची परिणती त्यांच्यातील परस्परसंबंध ठरविण्याच्या दृष्टीने अनेक गण, उपगण व कुलेही सुचविण्यात झाली आहेत. तथापि यांपैकी पुष्कळ जीवाश्मांचा काळ दीर्घ असल्याने स्तरवैज्ञानिक व खडकांचे वय ठरविण्याच्या दृष्टीने त्यांची उपयुक्तता थोडीच आहे मात्र पाणी व जमीन यांच्या स्थलांतराने परस्परसंबंधात झालेले बदल समजून घेण्यासाठी त्यांची मदत होते आणि पुराभूगोलाच्या दृष्टीने ते उपयुक्त आहेत. या संदर्भात भारतीय उपखंडाच्या दृष्टीने देता येणारे उदाहरण म्हणजे गोड्या पाण्यात राहणाऱ्या पश्चिम भारतामधील माशांच्या मलेशिया प्रदेशात असलेल्या माशांशी असणाऱ्या आप्तभावाविषयीचे एस्.एल्.व्होरा यांचे संशोधनकार्य हे होय.
पुष्कळ विविधता व मर्यादित स्तरवैज्ञानिक उपयुक्तता यांमुळे भारतातील अस्थिमत्स्यांचा विचार केवळ स्थूलपणेच करायला पाहिजे. भारतातील शैलसमूहांत आढळणाऱ्या अस्थिमत्स्यांच्या कित्येक गणांपैकी क्लुपिफॉर्मीस, पर्सिफॉर्मीस व सायप्रिनिफॉर्मीस गण लक्षात घेण्यासारखे आहेत.
क्लुपिफॉर्मीस गणाचे बहुतेक जीवाश्म मध्य प्रदेश, प्राणहिता-गोदावरी खोरे व ईशान्य सौराष्ट्रातील बध्वान क्षेत्र या भागांतील उत्तर क्रिटेशस कालीन अंतरा-ट्रॅपी थरांत आढळतात. तेथे क्लुपिया, होराक्लुपिया व म्यूस्पेरिया या वंशांचे जीवाश्म सापडले आहेत. दक्षिण भारतातील उत्तर क्रिटेशस कालीन अरियालूर थरांत एंकोडस वंशाचे, बिहारमधील राजमहाल टेकड्यांमधील उत्तर जुरासिक कालीन राजमहाल थरांत झिंग्रानिया वंशाचे आणि राजस्थानाच्या बारमेर जिल्ह्यातील कापुर्डीच्या मुलतानी मातीमध्ये स्क्रोंबोक्लुपिया, लैओमेनी व कापुर्डिया वंशांचे जीवाश्म आढळले आहेत. दक्षिण ट्रॅप खडकांतील अंतरा-ट्रॅपी थरांत पर्सिफॉर्मीस गणाचे यापेक्षाही जास्त जीवाश्म आढळले असून त्यांमध्ये इओसेरॅनस, नांदुस, पर्सिया, प्रिस्टोलेपिस व स्क्लॅरोपाजेस या वंशांचे जीवाश्म आहेत. आंध्र प्रदेशामध्ये राजमहेंद्रीजवळील काटेरू अंतरा-ट्रॅपी थरांमध्ये नांदुस व प्रिस्टोलेपिस या वंशांचे जीवाश्म सापडले आहेत. पॉलिमायेस वंशांचे जीवाश्म कापुर्डीजवळील इओसीन कालीन मुलतानी मातीत, तर त्रिचियुरस वंशांचे जीवाश्म बालासोरनजीकच्या मायोसीन कालीन खडकांत आढळले आहेत. पंजाबातील प्लायोसीन कालीन शिवालिक खडकांमध्ये सायप्रिनिफॉर्मीस गणाचे कित्येक जीवाश्म आढळले असून तेथे टॅचिसुरस, सिल्युरस, क्रायसिक्थिस, मायस्टा, रिटा, वॅगारियस, क्लॅरियस, हेटरोब्रँकस व सायप्रिनोडोंट हे वंश सापडले आहेत. टॅचिसुरस वंशांचे जीवाश्म बालासोर येथील मायोसीन थरांतही आढळले आहेत. काश्मिरातील प्लाइस्टोसीन कालीन कारेवा शैलसमूहात शिझोथोरॅक्स वंशांचे जीवाश्म सापडले आहेत. टेलिऑस्टिआय उपवर्गातील इतर गणांचे जीवाश्म भारतातील खडकांत विरळाच आढळतात. उदा., ऑफिसेफॅलिफॉर्मीस गणातील ऑफिसेफॅलस वंशांचे जीवाश्म पंजाबातील प्लायोसीन कालीन शिवालिक शैलसमूहात सापडले आहेत.
उभयचर वर्ग : [⟶ उभयचर वर्ग]. आधुनिक उभयचर (बेडूक, भेक, सॅलॅमँडर, न्यूट व सीसिलियन) अनियततापी (ज्यांच्या शरीराचे तापमान परिसराच्या तापमानावर अवलंबून असते असे) प्राणी आहेत. त्यांचे आयुष्य पाण्यात सुरू होते व नंतर ते तात्पुरते वा ठराविक काळाने जमिनीवर येऊन राहतात. यांचे माहीत असलेले सर्वांत पुरातन जीवाश्म म्हणजे पेनसिल्व्हेनिया (अमेरिका) भागातील उत्तर डेव्होनियन कालीन खडकांतील थिनोपस अँटिक्वसच्या सांगाड्याचे अवशेष आणि ग्रीनलंडमधील याच काळातील खडकांमधील इक्थिओस्टेगा व इक्थिओस्टेगॉप्सिस या वंशांतील प्राण्यांच्या कवट्या हे होत. हे जीवाश्म स्टेगोसेफॅलिया गणातील आहे, असे मानले जाते. थिनोपस अँटिक्वसमध्ये दोन पूर्णपणे वाढलेली बोटे आढळत असून तिसरे अल्पविकसित व चवथे त्याच्या अस्तित्वाचे केवळ सूचक इतपतच आढळते. अशा प्रकारे स्टेगोसेफॅलिया गणातील उभयचर हे जमिनीवरील पहिले पृष्ठवंशी प्राणी होत. त्यांच्यावरून असेही दिसते की, जमिनीवरील पृष्ठवंशींच्या पाच बोटे असलेल्या वैशिष्ट्यपूर्ण पायाचा पूर्वगामी म्हणजे या उभयचरांचा पाय होय.
दातांची रचना व कवटी यांत जटिलता येत जाण्याची प्रवृत्ती, गालावर अस्थिपट्टांचे आच्छादन, डोळ्यांभोवतीची श्वेत पटले, वल्ह्याप्रमाणे असलेले अंसपक्ष्यासारखे पाद यांवरून उभयचरांचे पूर्वज क्रॉसोप्टेरिजीयन मासे असावेत, असे सूचित होते.
(१) स्टेगोसेफॅलिया गण : यांना थोडीफार लांब शेपटी असते, कवटीवर अस्थिपट्ट असतात, शरीरावर त्वचीय खवले असतात किंवा शरीराच्या अधर भागावर व पुष्कळदा पृष्ठीन भागावर प्रशल्क (बाहेरचे खवले ) असतात. यांना जवळजवळ सारख्या लांबीचे पण आखूड दोन पाय असून पुढच्या पायांना चार तर मागच्यांना पाच बोटे असतात गोर्द (अवयवाच्या आतील स्पंजासारखे मऊ ऊतक) पोकळी रिकामी असलेले शंक्वाकार दात व दंतिनाला आतून घड्या पडून दातांना अधिकाधिक जटिल स्वरूप येण्याच्या प्रवृत्तीमुळे शेवटी जवळजवळ सर्व गोर्द पोकळी नष्ट होते व अशा दाताला लॅबिरिंथीय दात म्हणतात. ट्रायासिक कालीन लॅबिरिंथोडोंटांमध्ये असे आदर्श दात आढळतात. आधारक अस्थींवर किंवा कूपिकांमध्ये (खोबणीत) दातांचे रोपण झालेले असते. मणके पुष्कळदा आदिम प्रकारचे असून त्यांतील विविध भाग विविध प्रमाणात अस्थीभूत झालेले असतात.
सुरुवातीचे स्टेगोसेफॅलिया प्राणी आधुनिक सॅलॅमँडरप्रमाणे लहान होते. काहीतर त्याहूनही लहान होते, उदा., फायलोस्पॉंडायली उपगणातील मेलॅनर्पेटॉन वंशातील काही जातींच्या व लेपोस्पॉंडिलाय उपगणातील सीलिया प्युसिलाच्या पूर्व पर्मियन कालीन प्राण्यांची लांबी ३ सेंमी. पेक्षा कमी होती परंतु पुष्कळांचे आकारमान मोठे होते, उदा., २० ते ३० सेंमी.लांबीच्या कवट्या विरळा नव्हत्या. (टेम्नोस्पाँडायली उपगणातील एरिऑप्स मेगेसेफॅलस या न्यू मेक्सिको, टेक्सस इ. प्रदेशांतील पर्मियन कालीन खडकांत आढळलेल्या जीवाश्माची कवटी ४० ते ६० सेंमी.लांब व ३०-४६ सेंमी. रुंद होती, तर लॅबिर्रिथोडोंशिया गणातील मॅस्टोडॉनसॉरस या ट्रायासिक कालीन जीवाश्माची कवटी १.२५ मी.लांब होती आणि टेम्नोस्पाँडायलीच्या यूरॅनोसेंट्राडॉन सेनेकालेन्सिस या दक्षिण आफ्रिकेच्या ट्रायासिक कालीन जीवाश्मांची लांबी ३.७५ मी.होती).
उभयचरांचे जीवाश्मरूप अवशेष विपुल असले, तरी पुष्कळदा ते तुकड्यांच्या रूपात आढळतात व कवट्या, पायांच्या हाडासारखे सांगाड्याचे घटक, सुटे अथवा गटांनी आढळणारे मणके व दात यांसारख्या शरीरापासून अलग झालेल्या अवयवांचे जीवाश्म आढळतात. यांच्या पावलांचे ठसेसुद्धा विविध भागांत सापडले आहेत. पुष्कळ वंशांतील प्राण्यांच्या कवट्यांची तपशीलवार माहिती झालेली आहे व त्यामुळे अनेक बाबतींत सांगाड्यांची पुनर्रचना करणे शक्य झाले आहे. एरिऑप्स, ब्रॅंकिओसॉरस व आर्किगोसॉरस हे पुराजीव काळातील व मॅस्टोडॉनसॉरस हा ट्रायासिक कालीन उभयचरांचा वंश चांगला माहीत असलेल्या वंशापैकी आहे.
स्टेगोसेफॅलियाच्या जीवाश्मांच्या आढळण्याच्या तऱ्हांवरून ते सामान्यतः जमिनीवर व गोड्या पाण्याजवळ व काही सागरात राहत होते, असे दिसून येते. कार्बॉनिफेरस व पर्मियन काळांतील काही उभयचर दलदलीत रहात होते. स्टेगोसेफॅलियातील अधिक मोठे प्राणी बहुतकरून परभक्षी होते व त्यांची उपजीविका अधिक लहान उभयचर, मासे व कवचधारी प्राण्यांवर चालत असावी.
स्टेगोसेफॅलिया हा उभयचरांपैकी प्रमुख व पुराजीवविज्ञानाच्या दृष्टीने सर्वांत महत्त्वाचा गट आहे. त्यांचा काळ उत्तर डेव्होनियन ते ट्रायासिक अखेरपर्यंत असा आहे परंतु कार्बॉनिफेरस व पर्मियन काळांत ते सर्वाधिक विपुल व विविध प्रकारचे होते, तेव्हा पुष्कळांचे आकारमान मोठे झाले होते. खडकांच्या थरांचे सहसंबंध ठरविण्यासाठी उपयुक्त म्हणता येतील असे अधिक महत्त्वाचे वंश पुढील होत: आर्किगोसॉरस व ब्रँकिओसॉरस व त्यांच्याशी संबंधित कार्बॉनिफेरस व पर्मियन कालीन प्रकार, तसेच कॅपिटोसॉरस, मेटॉपोसॉरस, ब्रॅकिऑप्स व मॅस्टोडॉनसॉरस आणि त्यांच्याशी संबंधित ट्रायासिक कालीन प्रकार. एकेकाळी प्राचीन गोंडवनभूमीचे घटक असलेल्या व आता एकमेकांपासून पुष्कळ दूर गेलेल्या दक्षिण अमेरिका, दक्षिण आफ्रिका व भारत या प्रदेशांमधील यांचे जीवाश्म विशेष महत्त्वाचे आहेत.
भारतामध्ये गोंडवनी काळातील पृष्ठवंशी जीवाश्मांमध्ये स्टेगोसेफॅलियांचे जीवाश्म अतिशय महत्त्वाचे आहेत. काश्मीरमधील पर्मियन कालीन गँगॅमोप्टेरीस थरांत ॲक्टिनोडॉन, चेलिडोसॉरस, आर्किगोसॉरस व लायसेप्टरिजियम हे वंश सापडले आहेत तर बिहारमधील दगडी कोळशाच्या क्षेत्रातील पूर्व ट्रायासिक कालीन पांचेट थरांत गोनोओग्लिप्टस, ग्लिप्टोग्नॅथस, पॅचिगोनिया व पॅचिग्नॅथस या वंशाचे जीवाश्म सापडले आहेत. मध्य प्रदेशातील पूर्व ट्रायासिक कालीन बिजोरी थरांमध्ये गोंडवनसॉरस, लॅबिरिंथोडॉन व ऱ्हिनोसॉरस तर गोदावरी खोऱ्यातील पूर्व ट्रायासिक कालीन मांगळी थरांत ब्रँकिऑप्स वंशाचे जीवाश्म आढलले आहेत. गोदावरी खोऱ्यातील मध्य ट्रायासिक कालीन येर्रापल्ली थरांत पॅराटोसॉरस, उत्तर ट्रायासिक कालीन मालेरी थरांमध्ये पॅचिगोनिया, पँचिग्नॅथस व मेटॉपोसॉरस हे वंश सापडले आहेत. मध्य भारतातील उत्तर ट्रायासिक कालीन टिकी थरांतही मेटॉपोसॉरस या वंशाचे जीवाश्म आढळले आहेत. तसेच मध्य प्रदेशाच्या पंचमढी भागातील उत्तर ट्रायासिक कालीन बाग्रा व देन्वा थरांत मेटॉपोसॉरस वंश आढळला आहे.
या उभयचरांचे दात पोकळ शंक्वाकार प्रकारचे होते व दंतिनाला आतून घडया पडून लॅबिरिंथोडोंट प्रकार बनण्याची प्रवृत्ती आढळते. लॅबिरिंथोडोंट प्रकारचे दात उत्तर पर्मियन काळातील खडकांत सापडतात पण ट्रायासिक काळात अशा प्रकारचे दात प्रमुख होते. टेम्नोस्पाँडायली उपगणातील प्राण्यांच्या मणक्यांचे अस्थीभवन कमीअधिक प्रमाणात झालेले होते. स्टिरिओस्पाँडायलीमध्ये दोन्ही बाजूंनी किंचित अंतर्गोल चकत्या अस्थीभूत झालेल्या होत्या. लेपिडोस्पाँडायलीमध्ये डमरूसारखा छेद असलेले दंडगोलाकार, फायलोस्पाँडायलीमध्ये अपूर्ण पिंपाकार, तर ॲडेलोस्पाँडायलीमध्ये खोलवर दोन्ही बाजूंनी अंतर्गोल अशा दंडगोलाकार रूपात अस्थीभवन झालेले होते.
कवटीच्या घटकांच्या अस्थीभवनाच्या विविध पायऱ्या, मणक्यांचे स्वरूप व दात यांनुसार स्टेगोसेफॅलियांचे वर्गीकरण केलेले असते. कवटीच्या घटकांच्या अस्थीभवनाचा मुद्दा बाजूला ठेवून थोडक्यात पुढील विविध उपगण देता येतील. (क) टेम्नोस्पाँडायली: यात मणके कित्येक घटकांचे बनलेले असून त्यांतील काही सांधलेले असतात. दातांची प्रवृत्ती लॅबिरिंथीय प्रकाराकडे असते व काळ कार्बॉनिफेरस ते ट्रायासिक आहे. (ख) स्टिरिओस्पाँडायली: मणके पूर्णपणे अस्थीभूत चकत्या असतात व दोन्ही बाजूंनी किंचित अंतर्गोल असतात, दात लॅबिरिंथीय प्रकारचे व काळ ट्रायासिक आहे. (ग) लेपिडोस्पाँडीलाय : मणके दंडगोलाकार व छेद डमरूच्या आकाराचा व तंत्रिका-कमानी कशेरुक-कायाला सांधलेल्या असून दात साधे, पोकळ शंक्वाकार आणि काळ कार्बॉनिफेरस ते पर्मियन आहे. (घ) फायलोस्पाँडायली: मणके पिंपाच्या आकाराचे, अपूर्ण असून त्यांना तंत्रिका-कमानी व अधःकशेरुक-मूल (तंत्रिका-रज्जूच्या खालून निघणारी व कशेरुक-कायाचा भाग बनलेली उपास्थी)असतात, दात साधे, शंक्वाकार व पोकळ आणि काळ कार्बॉनिफेरस ते पर्मियन आहे. (च) ॲडेलोस्पाँडायली: मणके दंडगोलाकार, चांगले अंतर्गोलाकार दात साधे, पोकळ व बोथट शंकूच्या आकाराचे आणि काळ कार्बॉनिफेरस ते पर्मियन आहे.
फार पूर्वीच ई. डी. कोप यांनी पुढील गोष्ट सुचविली होती. टेम्नोस्पाँडायली उपगणाच्या स्टेगोसेफॅलियांचे कॉटिलोसॉरियांशी इतके निकटचे साम्य आहे की, ते सरीसृप वर्गाचे प्रत्यक्ष पूर्वज म्हणता येऊ शकतील. यांच्यातील दुहेरी पश्चकपालास्थि रुंद व कवटीची इतर वैशिष्ट्ये ही पुष्कळशी आधुनिक यूरोडेला व ॲन्यूरा यांच्या तशाच वैशिष्ट्यांसारखी आहेत.
(२) ॲन्यूरा गण : (सॅलेन्शिया, बॅट्रॅकिया, ईकॉडेटा). या गणात बेडूक व भेक येतात. याचा सर्वांत जुना जीवाश्म वायोमिंग (अमेरिका) येथील उत्तर जुरासिक कालीन इओबॅट्रॅकस व कदाचित स्पेनमधील याच काळातील मॉटेकोबॅट्रॅकस हा आहे. क्रिटेशस व नवजीव काळांतही अँन्यूरांचे जीवाश्म थोडेच आढळतात. राना, ब्यूफो, पॅलिओबॅट्रॅकस व इतर थोडे नवजीव काळातील जीवाश्म आहेत. प्लायोसीन आणि प्लाइस्टोसीन काळातील बहुतेक वंश आताही हयात आहेत. मुंबई भागातील क्रिटेशस (किंवा इओसीन प्रारंभीच्या) काळातील अंतरा-ट्रॅपी थरांत (बेडूक थरांत) इंडोबॅट्रॅकस वंशाचे जीवाश्म सापडले आहेत. अपूर्णावस्थेत टिकून राहिलेले व तुटपुंजे जीवाश्म यांमुळे उभयचरांच्या जीवाश्मांचा स्तरवैज्ञानिक सहसंबंध ठरविण्यासाठी फारच थोडा उपयोग होतो. जुरासिक काळाच्या आधीचे जीवाश्म नसल्याने उभयचरांची पूर्वजपरंपरा शोधणे कठीण झाले आहे. तथापि कवटीच्या आणि सांगाड्यातील घटकांच्या अस्थीभवनामध्ये घट होत जाण्याच्या प्रवृत्तीवरून अँन्यूरांचे संभाव्य पूर्वज पर्मियन काळातील ब्रँकिओसॉरससारखे उभयचर असावेत, असा कयास आहे.
सामान्य प्रकारचे पाय असलेल्या ॲन्यूरांच्या पूर्वजांची आणि अँन्यूरांमधील अशी ठळक वैशिष्ट्ये पुढील होत: अँन्यूरांचे मागील पाय अतिशय लांब असून मागील पायातील बाह्य जंघास्थी व अंतर्जंघास्थी आणि पुढच्या पायातील अरास्थी व प्रबाहु-अंतरास्थी या सांधल्या जाऊन त्यांच्या हालचालीच्या तऱ्हेला अनुकूल असा एक घटक बनलेला असतो आणि आखूड फासळ्यांच्या जागी आडवे प्रवर्ध (वाढी) हे दुसऱ्या दृष्टीने विशेषीकरण आहे. संरचनांच्या विविध घटकांच्या अस्थीभवनातील घट ही या दृष्टीने ऱ्हासाची प्रवृत्ती दर्शविते.
(३)यूरोडेला गण : (कॉडेटा). या गणात सॅलॅमँडर व न्यूट येतात. या गणाचा सर्वांत जुना जीवाश्म म्हणजे बेल्जियममधील पूर्व क्रिटेशस कालीन हायलीओबॅट्रॅकस क्रॉयी हा होय. लहान आकारमान व आकार, लांब शेपटी आणि जवळजवळ सारख्या व आखूड पायांच्या दोन जोड्या ही पूर्वजांची वैशिष्ट्ये (उदा., पुराजीव काळातील स्टेगोसेफॅलिया) या उभयचरांत बाह्यतः टिकून राहिलेली दिसतात परंतु शरीरांतर्गत सांगाड्याच्या कित्येक घटकांच्या अस्थीभवनातील घट झालेली दिसून येते व पुष्कळ घटक उपास्थिमय असतात. सामान्यतः यूरोडेला उभयचर लहान असतात परंतु बाडेन भागातील मायोसीन कालीन अँड्रिअस शुच्झेरी सु. १ मी. लांब होता. उत्तर पुराजीव काळातील अँडेलोस्पाँडायली प्राणी हे बहुतकरून यूरोडेलांचे पूर्वज असावेत. यूरोडेलांचे जीवाश्म अतिशय विरळाच व अपूर्णावस्थेत आढळतात. यांचे मध्यजीव कालीन केवळ तीनच सांगाडे माहीत असून नवजीव कालीन निक्षेपांत आढळलेले थोडे वंश हे आत्ताच्या वंशांपासून वेगळे आहेत असे समजणे अवघड आहे. यांचे जीवाश्म गोड्या पाण्यात साचलेल्या खडकांत मिळाले आहेत.
यूरोडेलांची उघडी त्वचा, भरीव मणके व अस्थीभवनातील घट ही मुख्यत्वे यूरोडेला व स्टेगोसेफॅलिया यांच्यातील भेद दर्शविणारी वैशिष्ट्ये आहेत.
(४)जिम्नोफॉयोना गण : (ॲपोडा).कृमिरूप पादहीन अशा उभयचर प्राण्यांचा हा लहान गट दक्षिण अमेरिका, दक्षिण आफ्रिका व भारत यांच्या उष्ण कटिबंधीय भागांत आढळतो. या गटाचे जीवाश्म अद्यापि आढळलेले नाहीत परंतु हे प्राणी पर्मियन काळातील लहान आकारमानाच्या लायसोरोफससारख्या ॲडेलोस्पॉँडायलस उभयचरांचे वंशज असू शकतील.
सरीसृप वर्ग : [ सरपटणाऱ्या प्राण्यांचा वर्ग ⟶ सरीसृप वर्ग]. सरीसृप अनियततापी असून त्यांचे श्वसन फुफ्फुसाद्वारे चालते आणि त्यांचा सर्व सांगाडा अस्थिमय असतो. साप व काही सरडे वगळता त्यांचा सर्व सांगाडा अस्थिमय असतो. साप व काही सरडे वगळता त्यांना पायांच्या दोन जोड्या असतात. बहुतेकांना शंक्वाकार दात असतात प्रगत किलोनिया व सरीसृपांच्या इतर गटांतील थोड्या तुरळक प्राण्यांना दात नसतात. त्वचा अनावृत्त असते अथवा तिच्यावर त्वचीय चिलखत (आवरण) किंवा खवले असतात. कवटीला बहुतकरून एक आधार (पश्चकपालास्थि-कंद) असतो.
नामशेष झालेले सरीसृप हे एकंदर सरीसृप वर्गाचा मोठा भाग

असून जीवाश्मांत संरचनात्मक विविधता बऱ्याच मोठ्या प्रमाणात दिसून येते.सरीसृपांचे बहुसंख्य जीवाश्म जमिनीवरील प्राण्यांचे आहेत. काही जलचर आहेत, तर काही उडणारे असून या जीवनप्रणालीला अनुसरून त्यांच्या पायांमध्ये परिवर्तन झालेले आहे.
सरीसृप वर्ग पर्मियन काळात अवतरले व बहुतेक मध्यजीव काळात ते पृष्ठवंशींपैकी प्रमुख प्राणी होते. त्यांच्यापैकी बहुतेक क्रिटेशस अखेरीस निर्वंश झाले आणि ॲलिगेटर, साप, मगरी व सरडे यांसारखे थोडेच प्राणी आता हयात आहेत. एच्.डी.ब्लॅंव्हील यांनी १८१६ साली व मेरेम यांनी १८२० साली उभयचरांपासून सरीसृप हा स्वतंत्र वर्ग म्हणून वेगळा केला आणि आर्.ओएन, टी.एच्. हक्सली, ओ. सी. मार्श, ई. डी. कोप, एच्. व्हॅन मायर आणि इतरांच्या संशोधनामुळे सरीसृपांचे समाधानकारक वर्गीकरण करणे शक्य झाले.
कानशीलाच्या (शंखक) भागाच्या विकासानुसार सरीसृपांचे ॲनॅप्सिडा, सिनॅप्सिडा, पॅरॅप्सिडा, डायप्सिडा व थेरॅप्सिडा असे गट पाडले जातात. विविध गणांच्या जातिवृत्त-संबंधाचे प्रतिबिंब या गटांत पडलेले दिसते.
कानशीलाचे, तसेच मणक्याचे स्वरूप, दात, हनुसंधिका (खालचा जबडा ज्याला जोडलेला असतो ते हाड), पाय, श्रोणि-कमान, वक्षीय कमान, त्वचेचे अस्थीभवन असणे किंवा नसणे इ. इतर बाबींची वैशिष्ट्ये यांचा एकत्र विचार करून सरीसृपांचे १४ (काहींच्या मते यापेक्षाही थोडे जास्त )गण पाडण्यात येतात.
(१) कॉटिलोसॉरिया गण: हे जमिनीवरचे सर्वांत आदिम सरीसृप आहेत. त्यांची कवटी पुष्कळदा दबलेली (खोलगट) असून तिचा माथा स्टेगोसेफॅलिया उभयचरांप्रमाणे सलग असतो. स्टेगोसेफॅलिया कॉटिलोसॉरियांचे जवळचे नातेवाईक असून त्यांच्याप्रणाणेच हेही मुख्यत्वे दलदलीत राहणारे होते. कॉटिलोसॉरियांचा काळ पूर्व पर्मियन ते ट्रायासिक असा असला, तरी त्यांचे जीवाश्म बहुतकरून दक्षिण आफ्रिका, यूरोप व उत्तर अमेरिका येथील पर्मियन कालीन खडकांतच अधिक आढळले आहेत.
(२) थेरोमॉर्फा गण : हे जमिनीवरील आदिम सरीसृप असून त्यांना आपल्या अवजड शरीराचे वजन तोलण्याइतके बळकट पाय होते मणके दोन्ही बाजूंनी अंतर्गोल होते श्रोणी व वक्षीय कमानीचे घटक भाग सायुज्यनाने किंवा, सीवनींच्या द्वारे घट्ट चिकटलेले असत. जबड्यांच्या कडांशी खळग्यांत असलेल्या दातांच्या ओळी असत दात कधीकधी पट्टांवर असत व काही प्रगत जातींमध्ये दातांचे विभेदन होण्याची प्रवृत्ती आढळते त्वचेचे अस्थीभवन झालेले नसे.
घटक हाडांच्या सायुज्यनाने बळकट झालेल्या श्रोणी व वक्षीय कमानीसारख्या सांगाड्याच्या घटक भागांच्या बाबतीत यांच्यात व सस्तन प्राण्यांत सर्वसाधारण सारखेपणा आढळतो व त्यावरून कोप यांनी या सरीसृपांना थेरोमॉर्फा असे नाव दिले आहे. याच्या थेरोडोंशिया या उपगणात दातांचे कृंतक, सुळे (रदनक), उपदाढा (अग्रचर्वणक) व दाढा (चर्वणक) असे विभेदन झालेले असून त्यांतील सायनोडोंशिया या उपकुलातील सायक्लोगॉंफोडॉन व पर्मोसायनोडॉन या वंशांमध्ये काही दातांची पुनःस्थापनाही झालेली आढळते. थेरोमॉर्फाचा काळ पूर्व पर्मियन ते ट्रायासिक अखेरपर्यंत असा आहे. दक्षिण आफ्रिका, भारत, रशिया, मध्य यूरोप, उत्तर अमेरिका, ब्राझील इ. भागांतील विशेषतः पर्मोट्रायासिक काळात या गणातील व त्यातल्या त्यात डिसिनोडोंशिया व थेरोडोंशिया उपगणातील वंशांच्या प्राण्यांमध्ये पुष्कळ विविधता आली होती, ते विपुल होते व त्यांचा विस्तृत प्रसार झाला होता.
थेरोमॉर्फा गणाचे पाच उपगण आहेत. त्यांपैकी पेलिकोसॉरिया हा सरड्यांसारख्या सरीसृपांचा छोटा गट आहे. त्याची कवटी लांब व रुंद व कधीकधी बाजूने दबलेली अशी असून त्यांची शेपटी पुष्कळदा लांब म्हणजे २-३ मी. लांब वाढलेली असे. डायमेट्रॉडॉन व टेनोसॉरसांप्रमाणे काहींमध्ये ठळक, साधा व बाजूने चापट झालेला तंत्रिका कंटक (मणक्याची काट्यासारखी वाढ)असे टेनोसॉरसच्या काही जातींमध्ये तो ६० सेंमी. पर्यंत लांब होता. या उपगणाचा कालावधी पूर्व पर्मियन ते ट्रायासिक असा आहे परंतु तो बहुतकरून पर्मियन काळातच होता. त्यांचे जीवाश्म मुख्यत्वे उ. अमेरिकेत व क्वचित यूरोपातही आढळले आहेत.
ड्रोमोसॉरिया उपगण हाही एक लहान गट होता. यातील प्राणी किरकोळ बांध्याचे, लहान आकारमानाचे होते त्यांचे पाय सडपातळ व शेपटी लांब असे. त्यांचे बहुतेक जीवाश्म दक्षिण आफ्रिकेतील पर्मियन कालीन खडकांत आढळले आहेत.
डायनोसेफॅलिया उपगणातील प्राण्यांचा बांधा थोराड होता व त्यांची लांबी २-३ मी. पर्यंत होती त्यांचे पाय काहीसे ताणलेले असत. टेपिनोसेफॅलिडी कुलातील प्राणी शाकाहारी, तर टिटँनोसुचिडी कुलातील प्राणी कदाचित मांसाहारी होते. या उपगणातील बहुतेक जीवाश्मही दक्षिण आफ्रिकेतील पर्मियन कालीन खडकांतच आढळले आहेत मात्र काही रशियातही सापडले आहेत.
डिसिनोडोंशिया उपगणातील प्राण्यांचे आकारमान घुशीपासून ते टॅपिर प्राण्याएवढे होते. त्यांचा बांधा थोराड असून डोके सापेक्षतः मोठे म्हणजे पुष्कळदा ५० ते ६० सेंमी. लांब असे. त्यांच्यापैकी पुष्कळ जमिनीवर राहणारे व लिस्ट्रोसॉरससारखे काही कदाचित जलचर होते. या उपगणात असंख्य वंश असून त्यांचा प्रसार विस्तृत प्रमाणात झालेला आहे. यांचा प्रसार विशेषतः दक्षिण आफ्रिका व भारत, ब्राझील यांसारख्या गोंडवनभूमीच्या प्रदेशांमध्ये पर्मियन व ट्रायासिक कालीन खडकांत झालेला असून रशिया, स्कॉटलंड व तुरळक प्रमाणात उत्तर अमेरिकेतही यांचे जीवाश्म आढळले आहेत. भारतामध्ये पूर्व ट्रायासिक कालीन पांचेत थरांत डिसिनोडॉन व लिस्ट्रोसॉरस तर मध्य ट्रायासिक कालीन येर्रापल्ली थरांत वाडियासॉरस व रेक्निसॉरस या वंशाचे जीवाश्म सापडले आहेत. एंडोथायोडॉन हा भारतातील सरीसृपांचा सर्वांत जुना जीवाश्म वंश असून तोही याच उपगणातील आहे. हे जीवाश्म प्राणहिता-गोदावरी खोऱ्यातील मध्य पर्सियन कालीन लोहमय ग्रंथिल (गाठाळ) वालुकाश्मात आढळले आहेत.
थेरिओडोंशिया या उपगणाचेही असंख्य वंश असून त्यांचे जीवाश्म दक्षिण आफ्रिका, भारत, ब्राझील, मध्य यूरोप, रशिया, उत्तर अमेरिका इ. भागांतील पर्मियन व ट्रायासिक कालीन खडकांत विस्तृत प्रमाणात आढळले आहेत. या उपगणाचे सर्वांत महत्त्वाचे वैशिष्ट्य म्हणजे रॉबर्ट ब्रूम (१८६६ – १९५१) यांनी इक्टिडोसॉरिया उपकुलात समाविष्ट केलेल्या या उपगणातील काही लहान आकारमानाच्या प्राण्यांचा गट हे होय. कवटीचे व सांगाड्याचे घटक भाग यांच्या बाबतीत या लहान प्राण्यांमध्ये व मोनोट्रिमॅटा या शिशुधान स्तनी प्राण्यांच्या गटामध्ये अतिशय साम्य असून हे दोन गट एकमेकांपासून वेगळे ओळखणे अतिशय अवघड असते. सायक्लोगॉंफोडॉन व पर्मोसायनोडॉन या वंशांमध्ये दातांचे कृंतक, सुळे, उपदाढा व दाढा असे विभेदन झालेले होते, तर स्कायलॅकोसॉरस वंशांमध्ये सुळ्यांच्या मागे दंतावकाश (दोन प्रकारच्या दातांमधील दंतहीन फट) निर्माण झाल्याचे आढळले आहे.
अशाप्रकारे थेरोमॉर्फा गणाचे एका बाजूने कॉटिलोसॉरियाकडून स्टेगोसेफॅलियाशी निकटचे संबंध आहेत, तर दुसऱ्या बाजूला थेरिओडोंशियाच्या इक्टिडोसॉरिया उपकुलातील प्राणी व मोनोट्रिमॅटा गटातील सस्तन प्राणी यांच्यात भेद करणे अवघड आहे. अशा तऱ्हेने थेरोमॉर्फा गण उभयचर व सस्तन प्राणी यांच्यामधील दुवा ठरतो.
(३)एरीओस्केलिडिया गण : लहान सरड्यासारख्या व जमिनीवर राहणाऱ्या सरीसृपांचा हा छोटा गट आहे. यातील प्राण्यांचे शरीर व पाय सडपातळ असून मागील व पुढील पाय जवळजवळ सारखेच लांब असतात. त्यांचे दात शंक्वाकार, खळग्यांत बसविलेले व काही टाळूवर असतात. या गणाचे थोडेच जीवाश्म स्वित्झर्लंड, फ्रान्स, इटली, जर्मनी, उत्तर अमेरिका इ.प्रदेशांतील पर्मियन व ट्रायासिक कालीन खडकांतच आढळले आहेत.
(४) मेसोसॉरिया गण : हा उभयचर वा जलचर आदिम सरीसृपांचा लहान गट आहे. यांची शेपटी बाजूने चापट झालेली व लांब असून कवटी लांब, अरुंद आणि मान आखूड होती. उभयचर प्रकारात मणके सडपातळ असून दात लांब, शंक्वाकार व जबड्याच्या कडांशी खळग्यांत बसविलेले व लहान दात पट्टावर असत. या सरीसृपांच्या कवटीच्या मागील भागांची पुरेशी माहिती उपलब्ध झाली नसल्याने त्यांचा गण असा स्वतंत्र दर्जा निश्चित झालेला नाही व ते इक्थिओसॉरिया, नोथोसॉरिया अथवा ऱ्हिंकोसेफॅलिया या इतर गणांपैकी असण्याची शक्यता आहे असे मानतात. मेसोसॉरिया ही कदाचित कॉटिलोसॉरियाची अगदी आधीची शाखा असावी आणि तिच्यातील प्राणी नंतर जलचर झाले असावेत. या गणाचे जीवाश्म केवळ गोंडवन भूमीतील (उदा.,दक्षिण आफ्रिका, भारत, ब्राझील इ.) पर्मियन कालीन खडकांतच आढळले आहेत.
(५) इक्थिओसॉरिया गण : (इक्थिओप्टेरिजिया गण). हा भूचरांची (जमिनीवर राहणाऱ्या प्राण्यांची) पूर्वजपरंपरा असलेल्या आदिम सागरी सरीसृपांचा गण आहे. त्यांना त्वचीय चिलखत नव्हते. त्यांचे शरीर माशांसारखे, शेपटी लांब होती व त्यांना जवळजवळ स्पष्ट अशी मान नव्हती. डोक्यावर चांगले विकसित झालेले निमुळते तुंड होते. त्यांची नेत्रकोटरे मोठी व श्वेत पटलांची कडी असलेली होती. त्यांना अणकुचीदार शंक्वाकार अनेक दात होते आणि दातांवर उभे चरे व दोन वा तीन कूटकही (उंचवटे) होते. ट्रायासिक प्रारंभीच्या काळातील प्राण्यांचे दात खळग्यात घट्ट बसविलेले होते परंतु नंतरच्या प्राण्यांचे दात सलग खोबणीत असत आणि मध्यजीव काळातील नंतरच्या काहींना दातच नव्हते. या प्राण्यांना असंख्य व दोन्ही बाजूंनी चांगले अंतर्गोल असलेले मणके होते. श्रोणी व वक्षीय कमानी बळकट होत्या. यांच्या पायांचे अनेक अंगुलास्थी असलेल्या वल्ह्यांसारख्या अवयवात परिवर्तन झालेले होते पायांची हाडे आखूड व बळकट असत. ते सागरात रहात असल्याने त्यांचे अवशेष चांगले टिकून राहीले व जवळजवळ पूर्ण असे पुष्कळ सांगाडे सापडले आहेत. या गणातील पुष्कळ प्राण्यांची लांबी १०-१२ मी. होती. यांच्या इंग्लंड व जर्मनीतील पूर्व जुरासिक कालीन काही सांगाड्यांच्या उदराच्या भागांत माखली व मासे या त्यांच्या भक्ष्यांचे जीवाश्मरूप झालेले अवशेष मिळाले आहेत. पूर्व जुरासिकमध्ये या गणाचे प्राणी विपुल होते व क्रिटेशस काळात त्यांचा विस्तृत प्रसार झाला होता, ते सागरात राहणारे असल्याने त्यांचा दोन्ही गोलार्धात विस्तृतपणे प्रसार झाला होता व या गणातील विशेषतः इक्थिओसॉरिडी कुलातील जीवाश्म विविध प्रदेशांतील खडकांच्या थरांचे सहसंबंध ठरविण्यासाठी पुष्कळ उपयुक्त आहेत. दक्षिण भारतामधील उत्तर क्रिटेशस कालीन अरियाळूर मालेच्या खडकांत या गणातील इक्थिओसॉरस वंशाचे जीवाश्म आढळले आहेत.
(६)प्लॅकोडोंशिया गण : हा जलचर अथवा उभयचर आदिम सरीसृपांचा छोटा गण आहे. यातील प्राण्यांना विशाल त्वचीय चिलखत, कमीअधिक लांब मुस्कट, आखूड मान, आखूड शेपटी व लांब युग्मित पाय होते. मात्र या गटाचे सर्वांत महत्त्वपूर्ण वैशिष्ट्य म्हणजे त्यांच्या टाळूवरील पट्टकीय स्वरूपाचे चुरडणारे दात, दंतिकास्थी (खालच्या जबड्यातील कला-अस्थी), अग्रजंभिका (जंभिकेच्या अग्रभागी असलेली हाडे) हे होय. त्यांना शंक्वाकार बळकट दातही असावेत. हे दातांचे पट्टकीय स्वरूप सरीसृपांमध्ये अतिशय अपूर्व असे आहे. या गणाचे जीवाश्म जर्मनी, स्वित्झर्लंड, सायलीशिया इ. प्रदेशातील पर्मियन व ट्रायासिक कालीन खडकांत आढळले आहेत.
(७) सॉरोप्टेरिजिया गण : या जलचर सरीसृपांना त्वचीय चिलखत नव्हते. त्यांची मान व डोके लांब आणि शेपटी आखूड होती पायांचे रूपांतर वल्ह्यांसारख्या अवयवात झालेले होते, मणके दोन्ही बाजूंनी चपटे होते व जबड्यांच्या कडांशी खळग्यात बसविलेल्या शंक्वाकार दातांच्या ओळी होत्या. लांब मान, आखूड शेपटी व वल्ह्यांसारख्या पायातील लांब हाडे यांमुळे या गणातील प्राण्यांच्या शरीराची ठेवणही सर्वसामान्य इक्थिओसॉरियातील प्राण्यांपेक्षा भिन्न होती. प्लेसिओसॉरस, पेलोन्यूस्टिस, नोथोसॉरस, इलॅस्मोसॉरस व सरड्यासारख्या प्राण्यांचा लॅरिओसॉरस या वंशाचे चांगले टिकून राहिलेले सांगाडे आढळले आहेत परंतु या गणातील इतरांचे जीवाश्म याहून कमी समाधानकारकपणे टिकून राहिलेले आढळतात. या गणाचे जीवाश्म जगातील पुष्कळ भागांत आढळले असून विशेषतः प्लेसिओसॉरिडी कुलाचे जीवाश्म विस्तृत प्रदेशांत सापडले आहेत म्हणून या कुलाचे जीवाश्म खडकांच्या थरांचे सहसंबंध ठरविण्याकरिता पुष्कळ उपयुक्त आहेत. या गणाचा काळ पूर्व ट्रायासिक ते उत्तर क्रिटेशस असा असून यांचे जीवाश्म जर्मनी, फ्रान्स, इटली, रशिया, इंग्लंड व उत्तर अमेरिका या प्रदेशांत आढळले आहेत. भारतामध्ये कच्छमधील उत्तर जुरासिक ते पूर्व क्रिटेशस कालीन उमिया मालेतील खडकांत या गणाच्या प्लेसिओसॉरस वंशाचे जीवाश्म सापडले आहेत.
एरीओस्केलिडिया गणातील ट्रॅनिस्टोफिअससारख्या भूचर वंशशाखेपासून बहुधा सॉरोप्टेरिजिया गणाचे प्राणी निर्माण झाले असून नंतर ते जलचर झाले असावेत. या गणातील काही प्राण्यांची लांबी फारच होती. उदा., इंग्लंडमधील पूर्व लायस कालीन ऱ्होमलिओसॉरस थॉर्नटोनी या प्राण्याची लांबी ७ मी. होती, तर प्लेसिओसॉरस व इलॅस्मोसॉरस वंशांतील काही जातींची लांबी १३ मी.च्या जवळपास होती. या गणातील प्राण्यांच्या काही सांगाड्यातील उदराच्या भागांत ज्यांवर त्यांची उपजीविका चाले अशा लहान सरीसृप, माखली व मृदुकाय प्राण्यांचे जीवाश्मरूप अवशेष आढळले आहेत. तसेच या उदर भागात खडे व गॅस्ट्रोलिथ (जठरातील कॅल्शियमी पुंज) म्हणून ओळखले जाणारे दगड हेही आढळले आहेत. त्यांवरून हल्लीच्या मगरीप्रमाणे हे प्राणी पचनास मदत व्हावी म्हणून दगड गिळत असावेत, असे दिसते.
(८)किलोनीया गण : या गणात कासवे व कूर्म यांचा समावेश असून त्यांच्यावरील कवचासारख्या आच्छादनामुळे ते इतर सरीसृपांपासून स्पष्टपणे वेगळे दिसतात. पृष्ठवर्म (कवचाचा पृष्ठीय भाग) हे पुढील पट्टांचे बनलेले असते. मध्यभागी अग्रपश्च दिशेतील आठ तंत्रिकीय वा कशेरुक पट्ट, अग्र टोकाजवळील रुंद षट्कोणी ग्रीवापृष्ठीय (मानेच्या मागील भागाचा) पट्ट व पश्च टोकाजवळील २ ते ४ पश्चतंत्रिकीय पट्ट. या मधल्या पट्टांच्या ओळीच्या दोन्ही बाजूंना पार्श्व वा परिफुप्फुसीय पट्टांच्या ८ अथवा काही सागरी जीवाश्मांमध्ये ९ किंवा १० आणि कडेलगतच्या सीमांत पट्टांच्या ११ वा १२ क्वचित १० जोडया असतात. कवचाच्या अधर भागामध्ये म्हणजे अधरवर्मामध्ये त्वचीय अस्थीभूत झालेले पट्ट यापेक्षा कमी असतात. त्यामुळे अग्रभागी डोके व पुढचे पाय आणि मागील भागात मागचे पाय व शेपटी बाहेर येण्याकरिता जागा रिकामी राहिलेली असते. उत्तर जर्मनीतील उत्तर ट्रायासिक कालीन ट्रायासोकीलिस या सर्वांत आधीच्या जीवाश्मांतही अशी रचना मूलतः निर्माण झालेली आढळते. यावरून किलोनिया गणाचा उदय त्याही पूर्वीच्या काळात झाला आहे, असे दिसते. हे प्राणी बहुतकरून मध्य पर्मियन कालीन कॉटिलोसॉरियांपैकी यूनोटोसॉरस सरीसृपांपासून आले असावेत कारण त्यांच्यातील मणक्यांचे स्वरूप अगदी किलोनियातील मणक्यांसारखे असल्याचे दिसून आले आहे. अगदी आधीचे काही प्राणी सोडता किलोनिया गणाच्या प्राण्यांना दात नाहीत पण त्यांच्या जबडयाच्या कडांवर पक्ष्यांच्या चोचीप्रमाणे शृंगी आवरण असते.
(८) किलोनिया गण : उत्तर ट्रायासिकमध्ये अवतरला असला, तरी उत्तर जुरासिक काळापासूनचे त्याचे जीवाश्म उत्तर अमेरिकेत व यूरोपात विस्तृतपणे आढळले आहेत पण पुष्कळदा जीवाश्म चांगले टिकून राहिलेले नसतात. क्रिटेशस व नवजीव काळातील पुष्कळ जीवाश्म आधुनिक वंशांशी चांगले निगडित आहेत व हे जीवाश्म चीन, ऑस्ट्रेलिया, उत्तर अमेरिका इ.प्रदेशांत आढळले आहेत. किलोनिया मुख्यत्वे जलचर असून काही थोडेच पूर्णतया सागरी आहेत व पुष्कळ भूचर आहेत. क्रिप्टोडायरा उपगणातील टेस्टयूडो गटाच्या काही कासवांचे आकारमान खूपच मोठे असून ती गालॅपागस, अँल्डॅब्रा, मॉरिशस, सिशल्स व रोंड्रीगेस या बेटांवर आहेत. मोठ्या आकारमानाच्या जीवाश्मांपैकी आर्किलॉन इश्विरास या डकोटातील (अमेरिका) क्रिटेशस कालीन जीवाश्मांची लांबी सु.२ मी. होती, तर पंजाबमधील शिवालिक शैलसमूहातील कोलोसोकीलिस ॲटलासची लांबी २ मी. पेक्षा जास्त होती.
निर्वंश झालेल्या आँफिकिलिडी या आदिम उपगणातील प्राण्यांना बहुतकरून आपले डोके व पाय कवचाच्या आत ओढून घेता येणे शक्य होत नव्हते प्ल्युरोडायरा उपगणाच्या प्राण्यांना डोके केवळ बाजूने आत घेता येत होते आणि बहुसंख्य जीवाश्म व सध्या राहात असलेले किलोनिया प्राणी ज्यात येतात त्या क्रिप्टोडायरा उपगणातील प्राणी आपले डोके सरळ कवचाच्या आत घेऊ शकतात.
किलोनियांच्या दीर्घकालीन इतिहासात त्यांच्या सर्वसाधारण रचनेत अल्पसेच परिवर्तन झालेले दिसते परंतु डर्मोकीलिस व ट्रिऑनिक्स या प्रथम महासागरात राहणाऱ्या आणि नंतर नद्यांमध्ये राहू लागलेल्या कासवांमध्ये सांगाड्याचे घटक लहान होऊन शरीराभोवती कातड्यासारखे आच्छादन विकसित झाले. हे आच्छादन बहुधा परिस्थितींशी जुळवून घेण्याकरिता निर्माण झाले असून काहीजण मानतात त्याप्रमाणे ते आदिम प्रकारचे वैशिष्ट्य नसावे. कधीकधी केवळ काही तंत्रिकीय पट्टच विकसित झालेले असतात आणि ऑस्ट्रेलियातील अलीकडच्या प्ल्युरोडायरांमध्ये तेही नाहीत.
मुंबई भागातील उत्तर क्रिटेशस (किंवा इओसीनच्या अगदी आधीच्या) काळातील अंतरा-ट्रॅपी थरांत (बेडूक थरांत) प्ल्युरोडायरामधील हायड्रॅस्पिस (प्लेटेमिस) हा जीवाश्म सापडला आहे. पंजाबातील प्लायोसिन कालीन उत्तर शिवालिक संघात किलोनियांचे कित्येक जीवाश्म आढळले आहेत. त्यापैकी हार्डेला, टेस्ट्यूडो, बेलिया, डॅमोनिया, जिओक्लेमिस, किलोन आणि कोलोसोकीलिस हे क्रिप्टोडायरा उपगणातील तर ट्रायोनिक्स, चित्रा व इमिडा हे ट्रायोनिकॉइडिया उपगणातील जीवाश्म आहेत.
(९)ऱ्हिंकोसेफॅलिया गण : हे सरड्यासारखे सरीसृप प्राणी आहेत. कधीकधी त्यांच्या अंगावर खवले असून मान आखूड असते. पाय काहीसे दुर्बल असून मागचे पाय पुढच्या पायांपेक्षा किंचित लांब असतात. त्यांचे मणके सामान्यतः दोन्ही बाजूंनी अंतर्गोल आणि नेत्रकोटरे व शंखक-रिक्ती (कवटीतील छिद्रे) काहीशा मोठ्या होत्या. दात अग्रदंती (हाडावरील उंचवट्याच्या माथ्याला सांधलेल्या) स्वरूपाचे असतात व ते जंभिका, अग्रहन, तालू व अधोहनू यांवर आणि क्वचित कूपिकेत असतात अथवा काहींना दात नसतातही.
मॅन्सफील्ड येथील उत्तर पर्मियन कालीन पॅलीओकॅमीलिओ हा लहान आकारमानाचा वंश बहुधा या गणाचा सर्वांत आधीचा वंश असावा. या वंशातील प्राण्यांचे अवशेष फारच अपूर्ण व विस्कळीत आहेत. तसेच सील्युरोसॉरॅव्हस हा मॅलॅगॅसीतील (मादागास्करमधील) पर्मियन कालीन वंशही या गणाचा असू शकेल. या दोन्ही वंशात कवटीच्या पश्च कडेशी त्रिकोणी कंटक होते. ऱ्हिंकोसॉरिडी कुल फक्त ट्रायासिक कालीन आहे, तर स्फीनोडोंटिडी कुलाचा काळ ट्रायासिक ते रीसेंट असा आहे. आता फक्त स्फीनोडॉन हा वंश न्यूझीलंडजवळील काही बेटांवरच आढळतो. या कुलांतील प्राण्यांचे वैशिष्ट्य म्हणजे आखूड व रुंद कवटी आणि तुंड चोचीप्रमाणे पुढे आलेले, हे होय. या दोन्ही कुलांतील पुष्कळ प्राणी उभयचर होते. ते पुष्कळ देशांतील ट्रायासिक कालीन निक्षेपांत सापडतात. यूरोप व गोंडवनभूमीच्या भारत, दक्षिण आफ्रिका, दक्षिण अमेरिका, मॅलॅगॅसी इ. प्रदेशांत हे जीवाश्म सापडले आहेत. भारतात गोदावरी खोऱ्यातील उत्तर ट्रायासिक कालीन मालेरी थरांमध्ये हायपेरॅडॅपेडॉन, पॅराडॅपेडॉन व पॅरासुचस या वंशाचे जीवाश्म आढळले आहेत. उत्तर अमेरिका, यूरोप व ईस्ट इंडीज या भागांतील उत्तर क्रिटेशस व इओसीन कालीन खडकांमध्ये कँपसोसॉरिडी कुलाचे जीवाश्म आढळले आहेत, तर थॅलॅटोसॉरिडीकुलाचे जीवाश्म बहुधा कॅलिफोर्नियातील केवळ ट्रायासिक कालीन खडकांतच सापडले आहेत. या दोन्ही कुलांतील प्राणी सागरांत राहणारे होते आणि त्यांची कवटी लांब व तुंड लांबोडके होते.
(१०) स्क्वॅमेटा गण : सरडे व सरडगुहिरे (लॅसर्टिलिया उपगण), साप व अजगर (ऑफिडिया उपगण) आणि जलचर सरड्यांचा निर्वंश गट (पायथॅनोमॉर्फा उपगण) हे या गणात येतात. त्यांचे मणके पश्चगर्ती (कशेरुक-काय मागील बाजूस अंतर्गोल असलेले) व क्वचित दोन्ही बाजूंनी अंतर्गोल असतात. शेपटी लांब असून ऑफीडिया वगळता इतरांना पाच बोटे असणाऱ्या पायांच्या दोन जोड्या असतात.
फ्रान्समधील उत्तर जुरासिक कालीन यूपोसॉरस, तसेच इंग्लंडमधील मॅसेलोडस ब्रॉडीइ व सॉरिलस ऑब्चूजस हे लॅसर्टीलियांचे सर्वांत पुरातन जीवाश्म आहेत. तथापि पर्मियन काळातील एरिओस्केलिस हा थेरोडोंशिया गणाचा सरडयासारखा प्राणी व स्क्वॅमेटा गणातील प्राणी यांच्यात निश्चित साम्ये आहेत. यावरून ऱ्हिंकोसेफॅलियाप्रमाणे स्क्वॅमेटा गण कदाचित पुराजीव कालीन कोणत्या तरी वंशजापासून आला असावा. अशा प्रकारे आणि स्क्वॅमेटा व थेरोडोंशिया हे सरीसृपांचे दोन गण आधीच एकमेकांपासून भिन्न होऊन गेले होते.
नवजीव काळातील लॅसर्टीलियांचे जीवाश्मही थोडेच असून ते बहुतेक आधुनिक कुलांतील म्हणता येतील, असे आहेत. हल्लीचा व्हॅरॅनस वंश प्लायोसीन काळापासून आहे. याचे उत्तर मायोसीन कालीन जीवाश्म पिकेर्मी(ग्रीस) येथे व भारतातील उत्तर शिवालिक संघात आढळले आहेत. पुष्कळ लॅसर्टीलियांचे आकारमान मोठे होते व सर्वांत मोठ्या मॅगॅलोनिया प्रिस्का या सरड्याची लांबी ५ मी.पेक्षा जास्त होती.
लॅसर्टीलियातील अर्ध-जलचर सरड्यांचे ऐगिॲलोसॉरिडी हे छोटे कुल फक्त पूर्व क्रिटेशस काळात आढळत असून त्यापासून पायथॅनोमॉर्फा हा सागरी सरडयांचा उपगण उदयास आला. या उपगणाचे जीवाश्म केवळ उत्तर क्रिटेशस काळातील खडकांत उत्तर अमेरिका, उत्तर आफ्रिका व यूरोपात विस्तृतपणे आढळतात. यातील पुष्कळ प्राणी लांब होते व त्यापैकी मोसॅसॉरस व टायलोसॉरस या वंशांतील प्राण्यांची कवटी जवळजवळ १ मी. लांब होती, तर प्लेटिकॉर्पस प्राण्यांची लांबी ५ मी. पेक्षा जास्त होती.
पॅचिऑप्सिस हा यूगोस्लाव्हियातील पूर्व क्रिटेशस कालीन जीवाश्म ऑफीडिया उपगणाचा सर्वांत जुना जीवाश्म असून तो कोलोफीडी या आदिम कुलातील आहे या कुलातील प्राणी बहुधा सागरात राहणारे होते आणि ते क्रिटेशस कालीन डोलोकोसॉरिडी कुलातील जलचर सरड्यांपासून सहजपणे वेगळे ओळखता येत नाहीत. या उपगणातील जीवाश्मांच्या नोंदी अतिशय विस्कळीत असून त्यांचे जीवाश्म बहुधा मणक्यांच्या रूपात आढळत असल्याने त्यांच्यापैकी पुष्कळांचे वर्गीकरणातील योग्य स्थान अनिश्चित आहे. भारतामध्ये पंजाबातील प्लायोसिक कालीन शिवालिक शैलसमूहात या उपगणातील पायथॉन मोल्युरस हा जीवाश्म सापडला आहे.
ऑफीडिया उपगणाचे बहुतेक प्लाइस्टोसीन कालीन प्राणी हे हल्ली असणाऱ्या वंशातील म्हणता येतील. आत्ताच्या ऑफीडियांप्रमाणे जीवाश्मरूप झालेले प्राणीही मुख्यत्वे भूचर, काही उभयचर व काही सागरी, काही बहुधा बिळांत राहणारे होते. (उदा.,अर्जेंटिनातील उत्तर क्रिटेशस कालीन डायनिलायसिस). विषारी सापांचा सर्वांत जुना जीवाश्म मॉसबाख-बायाब्रिख येथील मायोसीन कालीन प्रोव्हायपरा हा आहे. सागरातील काही साप फार लांब होते (उदा.,पॅलिऑफीडी या निर्वंश झालेल्या कुलातील साप) इटलीतील मॉंते बोल्का भागातील इओसीन कालीन आर्कॅऑफिस प्रोॲव्हस या सापाला ५६५ मणके होते तर ईजिप्तमधील मध्य इओसीन कालीन जायगँटो पायथन हा साप ९ मी.पेक्षा जास्त लांब होता.
पायथॅनोमॉर्फ प्राण्यांचा उत्तर क्रिटेशस काळात विस्तृत प्रमाणात प्रसार झाला होता, त्यामुळे या काळातील खडकांच्या थरांचे सहसंबंध ठरविण्यासाठी त्यांचा काहीसा उपयोग होतो. मात्र लॅसर्टीलिया व ऑफिडियांच्या जीवाश्मांच्या नोंदी अपूर्ण असल्याने त्यांचा उपयोग अत्यल्पच होतो.
(११) थीकोडोंशिया गण: ऱ्हिंकोसेफॅलियांप्रमाणे या गणातील प्राण्यांची शेपटी लांब, मान आखूड व पाय काहीसे दुर्बळ होते. पॅरासुचिया उपगण वगळता मुस्कट आखूड होते. त्यांना जबड्यांच्या कडांशी खळग्यांत रोवलेले दात असत. त्वचीय चिलखत चांगले विकसित झालेले होते. संरचनात्मक दृष्ट्या हा सरीसृपांचा सर्वसामान्यीकरण झालेला गट असून पॅरासुचिया हा उपगण क्रोकोडिलिया व डायनोसॉरिया गणांचा पूर्वज असावा.
स्यूडोसुकिया हे जमिनीवर राहणारे सरड्यांसारखे लहान प्राणी होते व त्यांची लांबी क्वचित १ मी. होती. त्यांचे जीवाश्म दक्षिण आफ्रिका, यूरोप, उत्तर अमेरिका व इंग्लंड या भागांतील ट्रायासिक निक्षेपांत आढळले आहेत. पेलिकोसिमिया उपगणातील प्राण्यांचे मुस्कट आखूड होते व त्यांना बाजूंनी दबलेले दंतुर सीमांत दात होते. या उपगणाचे जीवाश्म फक्त ट्रायासिक काळातील असून ते बहुतेक दक्षिण आफ्रिकेत सापडले आहेत पण अमेरिकेच्या संयुक्त संस्थानांतही या उपगणाचे जीवाश्म आढळतात. भारतात गोदावरी खोऱ्यातील उत्तर ट्रायासिक कालीन मालेरी थरांमध्ये या उपगणाच्या एरीथ्रोसुचस वंशाचे जीवाश्म सापडले आहेत.
ईओसुचिया उपगणाचे जीवाश्म पर्मियन व ट्रायासिक कालीन खडकांत मुख्यत्वे दक्षिण आफ्रिकेत आढळले आहेत. टँगॅसॉरिडी कुलातील प्राणी जलचर होते त्यांचे शरीर व मान आखूड आणि शेपटी लांब होती. यांचे जीवाश्म दक्षिण आफ्रिका व मॅलॅगॅसी येथील पर्मियन निक्षेपांत आढळले आहेत. यंगिनिडी कुलातील प्राण्यांचे मुस्कट दबलेले व सडपातळ असून कधीकधी कवटीवर शिल्पकाम केल्यासारख्या आकृती आढळतात. या कुलाचे जीवाश्म दक्षिण आफ्रिका व मॅलॅगॅसी येथील पर्मियन व ट्रायासिक कालीन खडकांत आढळले आहेत.
पॅरासुचिया गणात त्वचीय पट्ट चांगले विकसित झालेले होते व मुस्कट लांबोडके होते. बेलोडोंटिडी कुलात डोक्यावर शिल्पसदृश्य आकृती होत्या व ते केवळ ट्रायासिक काळातच आढळते. बेलोडॉन वंशाचा विस्तृत प्रसार झालेला असून त्याचा खडकांचे सहसंबंध ठरविण्यासाठी उपयोग होतो. डेस्मॅटोसुचिडी कुलातील अगदी थोड्याच प्राण्यांच्या शरीरावर त्वचीय अस्थिमय आच्छादन पसरलेले होते. या कुलाचे जीवाश्म केवळ ट्रायासिक काळातील खडकांत आढळले आहेत. भारतात या उपगणातील बेलोडॉन व ब्रँकिसुचस वंशाचे जीवाश्म गोदावरी खोऱ्यातील मालेरी थरांत सापडले आहेत.
(१२) क्रोकोडिलिया गण: क्रोकोडिलियांची नमुनेदार वक्षीय कमान सर्वांत आधीच्या क्रोकोडिलियांपैकी नोटोकँप्सा व पेडेटिकोसॉरस या वंशांतील प्राण्यांमध्ये आढळते व एरिथ्रोकँप्सामध्ये नमुनेदार श्रोणि-कमान सर्वप्रथम आढळली आहे. मात्र त्यांचे पाय सडपातळ असून ते जलचर होते. यांचे सर्व जीवाश्म दक्षिण आफ्रिकेतील पूर्व जुरासिक कालीन स्टॉर्मबर्ग थरांमध्ये आढळले आहेत. जुरासिक व पूर्व क्रिटेशस कालीन सर्व वंशांत मणके दोन्ही बाजूंनी अंतर्गोल किंवा दोन्ही टोके सपाट असलेले असतात. उत्तर क्रिटेशस व तृतीय कालीन जवळजवळ सर्व वंशांतील प्राण्यांचे अग्रत्रिकास्थीय (पृष्ठवंशाच्या टोकाच्या हाडाच्या आधीचे) मणके अग्रगर्ती (अग्रबाजू अंतर्गोल असलेले) असतात. पृष्ठीन चिलखत असल्यास ते प्रशल्कांचे बनलेले असते. प्रशल्कांवर बहुधा खळगे व क्वचित शिल्पाकृती आणि कणा असतो, त्यांच्या बहुतेक भागांवर नेहमी शृंगी बाह्य त्वचेचे आवरण असते. अधर बाजूवरील खवल्यांवर कमी शिल्पाकृती असतात व ते चापट असून त्यांना कणा नसतो. क्रोकोडिलियांना असंख्य शंक्वाकार दात असून दाताचा आडवा छेद लंबवर्तुळाकार किंवा वर्तुळाकार असतो. दातांवर चरे आणि कूटक वा खोबणीही असतात दात जबड्यांच्या कडांशी कूपिकांत रोवलेले असतात. एका जातीमध्ये व पुष्कळ वंशांमध्येही दातांची संख्या सामान्यतः स्थिर राहत असल्याचे दिसून येते. त्यांची मान आखूड असून पाय शरीराला आधार देण्याच्या दृष्टीने काहीसे कमजोर असतात.
क्रोकोडिलिया गण थीकोडोंशिया गणातील पॅरासुचियांपासून उदयास आल्याचे मानतात आणि क्रोकोडिलियाचे मेसोसुचिया व यूसुचिया असे दोन उपगण करतात.
मेसोसुचिया हा सर्वांत आधीचा उपगण पूर्व जुरासिक काळात अवतरला. यातील सर्वांत आधीच्या प्राण्यांचे तुंड लांब होते व उत्तर जुरासिकमध्येच आखूड व रुंद डोके असलेले वंश आढळतात. यांतील खुजे प्राणीही याच सुमारास अवतरले व त्यांचे हे अवतरणे बहुतकरून लहान सस्तन प्राण्यांच्या पृथ्वीवर अवतरण्याशी निगडित असावे. त्यांच्या पोटातील जीवाश्मरूप अवशेष व विष्टाश्म आणि त्यांची सर्वसामान्य शरीररचना यांवरून ते जलचर व बहुधा सागरात राहणारे होते, असे दिसते. बहुतेक जुरासिक काळात या प्राण्यांत अल्पच बदल झाला पण जुरासिकच्या शेवटी व क्रिटेशसच्या प्रारंभी त्यांनी गोड्या पाण्यात व नदीत राहण्याची जीवनप्रणाली अंगिकारल्याचे दिसून येते. कारण ते गोड्या पाण्यातील व भूचर प्राण्यांच्या, उदा., फोलिडोसॉरिडी कुलातील प्राण्यांच्या, जीवाश्मांबरोबर आढळले आहेत.
मेसोसुचियांचे मणके दोन्ही बाजूंनी अंतर्गोल किंवा दोन्ही टोके सपाट असलेले असे होते. टेलिओसॉरिडी, फोलिडोसॉरिडी व मेट्रिओऱ्हिंकिडी कुलांत तुंड लंबोडके होते मेट्रिओऱ्हिंकिडी कुलांतील प्राण्यांना त्वचीय चिलखत नव्हते. ॲटोपोसॉरिडी, गोनिओफॉलिडी व नोटोसुचिडी या कुलांतील प्राणी लहान सरड्याप्रमाणे होते व त्यांचे मुस्कट आखूड होते. नोटोसुचिडी कुलात त्वचीय चिलखत नव्हते व गोनिओफॉलिडी कुलामध्ये बळकट दंतुर कडा असलेले दात होते आणि पुष्कळदा त्यांच्यावर खोबणी असत.
पुष्कळ मेसोसुचियांची लांबी बरीच होती. उदा.,मॉंटॅनातील (अमेरिका) उत्तर क्रिटेशस कालीन टेलिओऱ्हिंकस ब्राऊनी, पेटरबरो येथील ऑक्सफर्ड क्लेमधील मिक्टेरोसुचस नॅसुटस व टयूनिसमधील पूर्व इओसीन काळातील डायरोसॉरस फॉस्फॅटिक्स प्राण्यांची कवटी १ मी. लांब होता, तर कॉनिग येथील मिस्ट्रिओसॉरस चॅपमनी प्राण्याची लांबी ६ मी. पेक्षा जास्त होती.
मेसोसुचियांचा काळ पूर्व जुरासिक ते उत्तर क्रिटेशस असा असून त्यांपैकी टेलिओसॉरिडी व फोलिडोसॉरिडी ही कुले कदाचित इओसीन काळामध्येही अस्तित्वात असावीत.
यूसुचिया उपगणात क्रिटेशसमधील बहुतेक, तृतीय काळातील व हल्लीचे क्रोकोडिलिया प्राणी येतात. त्यांचे अग्रत्रिकास्थीय मणके अग्रगर्ती असतात बहुतेकांचे मुस्कट रुंद असते. हायलीओचँप्सिडी हा लहान प्राण्यांचा आदिम गट केवळ पूर्व क्रिटेशस कालीन खडकांत आढळतो. गेव्हिॲलिडी कुलातील प्राण्यांचे तुंड लांब व निमुळते असून जबड्याच्या प्रत्येक बाजूला २५ ते ३५ दात असतात. भारत व ब्रह्मदेशातील तृतीय ते रीसेंट काळातील खडकांत या कुलांचे जीवाश्म आढळले आहेत. हे प्राणी पुष्कळच लांब होते, उदा., शिवालिक शैलसमूहात आढळलेल्या ऱ्हँपोसुचस कॅसिडेन्स या राक्षसी प्राण्याची लांबी १५ मी. होती. स्टोमॅटोसुचिडी या उत्तर क्रिटेशस कालीन कुलातील प्राण्यांना बहुतकरून पेलिकन पक्ष्यांप्रमाणे अधोहनु-कोष्ठ (पिशवीसारखी संरचना) होते. ईजिप्तमधील स्टोगॅडोसुचस इनर्मिस या प्राण्याची कवटी २ मी. लांब होती. क्रोकोडिलिडी व टोमिस्टोमिडी कुलातील प्राण्यांना अधर बाजूवर चिलखत नसते.
मसोसुचियांप्रमाणे यूसुचियांतही लांब व आखूड मुस्कट असलेले वंश आहेत. गेव्हीॲलिडी कुलाचा उदय आधुनिक प्रकारच्या मगरी व ॲलिगेटर यांपासून स्वतंत्रपणे झाला की, मेसोसुचिया या पूर्वजांपासून हे सर्व उत्पन्न झाले आहेत, हे निश्चित समजलेले नाही.
ॲलिगेटरिडी व क्रोकोडिलिडी या कुलांचे जीवाश्म यूरोप, दोन्ही अमेरिका, भारत व आफ्रिका या प्रदेशांतील गोड्या पाण्यात साचलेल्या उत्तर क्रिटेशस व तृतीय कालीन निक्षेपांत सापडले आहेत. दक्षिण अमेरिका, आफ्रिका, ऑस्ट्रेलिया, भारत, ईस्ट इंडिया, न्यू गिनी इ. भागांतील या कुलांतील अलीकडचे प्राणी हे फक्त उष्ण कटिबंधीय भागांतच आढळतात. भारतात पंजाबमधील प्लायोसीन कालीन उत्तर शिवालिक शैलसमूहांमध्ये यूसुचियां उपगणातील क्रोकोडिलिया, गेव्हिॲलिस व ऱ्हँपोसुचस या वंशांतील प्राण्याचे जीवाश्म सापडले आहेत.
(१३) डायनोसॉरिया गण : [⟶डायनोसॉर]. मध्यजीव काळातील सरीसृपांच्या या गणात अतिशय विविधता आढळते. पुष्कळ बाबतींत या गटाचे क्रोकोडिलिया व ऱ्हिंकोसेफॅलिया तसेच पक्ष्यांशीही पुष्कळ साम्य आहे (उदा., श्रोणि- कमान व मागच्या पायांच्या बाबतींत थेरोपोडा प्राण्यांचे यांच्याशी साम्य आहे).
डायनोसॉरांचे सर्वांत पुरातन जीवाश्म ट्रायासिक निक्षेपांतील असून त्यांच्या पायांची रचना व पावलांचे ठसे यांच्यावरून ते द्विपाद होते आणि त्यांच्या दातांवरून ते मांसाहारी होते, असे दिसून येते. तथापि सॉरोपोडा व ऑर्थोपोडा या उपगणांचे जुरासिक व क्रिटेशस काळांतील प्राणी शाकाहारी होते (कदाचित काही सर्वभक्षी होते). या दोन गणांतील काही द्विपाद काही चतुष्पाद प्राणी हे द्विपाद थेरोपोडा प्राणी पूर्वजांपासून उत्पन्न झाले असावेत.
डायनोसॉरांचे पुढील पाय मागील पायांपेक्षा सामान्यत अधिक आखूड तर काहींचे फारच आखूड असत आणि बोटे नखरांच्या वा खुराच्या आकाराची होती. मागच्या पायांना ३ वा ५ बोटे असत. त्यांची शेपटी व मान लांब असे. बहुतेक डायनोसॉरांची त्वचा अनावृत्त असे मात्र काहींच्या त्वचेवर एकमेकांवर न येणारे शृंगी खवले होते. थेरोपोडा व ऑर्थोपोडा यांतील काही प्राण्यांमध्ये अस्थिमय पट्ट, कंटक किंवा एकमेकांत गुंतलेले प्रशल्क यांचे बनलेले त्वचीय चिलखत धडाचा काही भाग व शेपटी यांच्यावर पसरलेले होते. मणके दोन्ही बाजूंनी सपाट किंवा पश्चगर्ती व क्वचित दोन्ही बाजूंनी अंतर्गोल असत. दात चमच्याच्या आकाराचे असून पुष्कळदा त्यांच्या पार्श्व कडा धारदार व दातेरीही असत. दात जबड्यांच्या कडांशी खळग्यांत किंवा खोबणीत रोवलेले असत.सर्व पृष्ठवंशी प्राण्यांत डायनोसॉरांची कवटी सामान्यत एकूण शरीराच्या मानाने लहान होती. ब्रॉंटोसॉरसची कवटी व ट्रायसेरॅटॉप्समधील मस्तिष्क – गुहा (मेंदूसाठी असलेल्या पोकळ्या) बहुधा सर्वांत लहान होत्या. जुरासिक व क्रिटेशस कालीन थेरोपोडा व ऑर्थोपोडांतील द्विपाद यांच्या कवटीची लांब बाजू मानेशी जवळजवळ काटकोनात होती परंतु ट्रायासिक कालीन थेरोपोडा, तसेच सॉरोपोडा व ऑर्थोपोडांतील चतुष्पाद यांच्या कवटीची लांब बाजू जवळजवळ कशेरुक- अक्षाच्या दिशेत होती.
सॉरोपोडा जीवाश्मांच्या उदराच्या भागांत खडे आढळले आहेत. सॉरोपोडा व ऑर्थोपोडा यांच्या सांगाड्याच्या अवशेषांबरोबर अंडी सापडली आहेत. या अंड्यांच्या कवचाची रचना आधुनिक सरीसृपांच्या अंड्यांच्या कवचाच्या रचनेपेक्षा वेगळी आहे पण या अंड्यांच्या कवचाची जाडी त्रिपार्श्विक स्तर (प्रचिनासारख्या कोशिकांचा बनलेला थर) ही आधुनिक पक्ष्यांच्या अंड्यांशी जुळणारी आहेत.
डायनोसॉरांचे जीवाश्म प्रथम इंग्लंडमधील उत्तर जुरासिक कालीन आणि वीडेन निक्षेपांत अठराव्या शतकाच्या प्रारंभी सापडले . बकलँड विल्यम यांनी १८२४ साली व जी. ए. मँटेल यांनी १८२५ साली ह्यांचे वर्णन केले. आर्. ओएन यांनी १८४१ साली डायनोसॉरिया गण प्रस्थापित केला. डायनोसॉरचा पहिला संपूर्ण सांगाडा इंग्लंडमधील वीडेन निक्षेपांत आढळला तो म्हणजे इग्वॅनोडॉन हा होय. जर्मनीतील ट्रायासिक कालीन पुष्कळ निक्षेपांत डायनोसॉरांच्या आदिम वंशांचे जीवाश्म आढळले आहेत. उत्तर अमेरिकेतील सर्व निक्षपांत डायनोसॉरांचे जीवाश्म आढळले आहेत.
डायनोसॉर ट्रायासिकमध्ये अवतरले, जुरासिक व क्रिटेशस काळांत त्यांचा विकास कळसाला पोहोचला आणि क्रिटेशसच्या अखेरीस ते निर्वंश झाले. त्यांचा प्रसार जगभर झालेला असल्याने निक्षेपांचे स्तरवैज्ञानिक सहसंबंध निश्चित करण्यासाठी डायनोसॉरांचा पुष्कळ उपयोग झाला आहे. यांचे जीवाश्म विशेषकरून दक्षिण आफ्रिका, मॅलॅगॅसी, भारत, चीन व ब्राझील या प्रदेशांत आणि बऱ्याच प्रमाणात यूरोप, उत्तर व दक्षिण अमेरिका या भागांतही आढळले आहेत. ऑस्ट्रेलियातील ट्रायासिक व क्रिटेशस कालीन खडकांत ते थोडेच आढळतात.
डायनोसॉरांत खूप विविधता आली होती. सामान्यतः त्यांचे आकारमान मोठे होते. त्यांच्यापैकी काहींचे, विशेषतः काही सॉरोपोडांचे, आकारमान राक्षसी होते व एवढे आकारमान इतर भूचर पृष्ठवंशी प्राण्यांत आढळत नाही.
ओ. सी. मार्श यांनी डायनोसॉरीयांचे पुढील तीन उपगण पाडले आहेत: (१) थेरोपोडा, (२) सॉरोपोडा आणि (३) प्रिडेंटॅटा. एच्. जी. सीली यांनी डायनोसॉरीयांचे पुढील दोन गट केले आहेत: (अ) सॉरिश्चिया (यात वरील थेरोपोडा व सॉरोपोडा येतात) व (आ) पुढील तीन उपगण ऑर्निथिशिया (हा गट ई. डी. कोप यांच्या ऑर्थोपोडाशी व मार्श यांच्या प्रिडेंटॅटाशी तुल्य आहे). ए. एस्. रोमर यांच्या मते ऑर्निथिशियाचे पुढील चार उपगण करता येतील: (१) ऑर्निथोपोडा, (२) स्टेगोसेफॅलिया, (३) अँकिलोसॉरिया व (४) सेरॅटॉप्सिया.
(अ) सॉरिश्चिया: यांच्यापैकी मांसाहारींचे दात शंक्वाकार, तीक्ष्ण, सुरीप्रमाणे चापट होते, तर शाकाहारींचे दात बोथट असत. मांसाहारींच्या बोटांना नख्या होत्या.
थेरोपोडा उपगणातील प्राण्यांचे मागचे पाय लांबट व मान लांब होती दात लांब, धारदार, चापट पुष्कळदा दंतुर कडांचे (करवतींसारखे ) व वाकडे होते म्हणजे असे दात मांसाहाराला सुयोग्य होते. आधीचे प्राणी लहान चणीचे होते व त्यांची हाडे पोकळ होती. अशा तऱ्हेने हल्क्या शरीरामुळे ते जलदपणे हालचाल करू शकत असत. ट्रायासिक काळातील आधीच्या प्राण्यांचे आदिम मणके दोन्ही बाजूंनी अंतर्गोल अथवा दोन्ही टोके सपाट असलेले असे होते परंतु जुरासिक व क्रिटेशस काळांतील प्राण्यांचे अग्रत्रिकास्थीय मणके अग्रगर्ती होते.
मोठ्या चणीचे प्राणीही दक्षिण आफ्रिका, यूरोप व उत्तर अमेरिका येथील उत्तर ट्रायासिक कालीन खडकांत आढळले आहेत आणि यांच्यापैकी काहींपासून मेगॅलोसॉरस, ॲल्लोसॉरस, सेरॅटोसॉरस इ. थोराड चणीचे वंश निर्माण झाले असावेत. यांची कवटी आधीच्या प्राण्यांच्या मानाने शरीरापेक्षा काहीशी मोठी होती. क्रिटेशसमध्ये मांसाहारीचे शरीर अधिकच अवजड झाले. त्यांचे पुढचे पाय आखूड व कमजोर होते. काहींचे उलट वळलेले करवतींसारखे दात सु. १५ सेंमी. लांब असत, उदा., टायरॅनोसॉरसचे काही नातेवाईक. या उपगणातील काही प्राणी प्रचंड आकारमानाचे होते. उदा., टायरॅनोसॉरस रेक्स १० मी.लांब होता. क्रिटेशसच्या अखेरीस हा उपगण निर्वंश झाला. थेरोपोडांचे पुष्कळ जीवाश्म भारतात आढळले आहेत. बिहारमधील दगडी कोळशाच्या क्षेत्रातील पांचेत थरांत एपिकँपोडॉन गोदावरी-प्राणहिता खोऱ्यातील ट्रायासिक कालीन मालेरी थरांत सील्युरोसॉरिया व मेसोस्पॉंडायलस, मध्यप्रदेशातील उत्तर क्रिटेशस कालीन लॅमेटा थरांत इंडोसुचस, इंडोसॉरस, जबलपोरिया, लॅमेटासॉरस, कॉंप्रोसॉरस, लीव्हिसॉरस, सील्युरॉइड, ड्रिप्टोसॉरॉइड, ऑर्निथोमिमॉइड व मेसोस्पॉंडायलस या वंशाचे तर उत्तर क्रिटेशस कालीन अरियालूर मालेत मेगॅलोसॉरस वंशाचे जीवाश्म सापडले आहेत. मध्य भारतातील टिकी थरांत मेसोस्पॉंडायलसचे जीवाश्म आढळले आहेत.
क्रिटेशस काळात काही शहामृगासारखे थेरोपोडा होते, उदा., उत्तर अमेरिकेतील स्ट्रुथिओमायमस व ऑर्निथिओमायमस यांचे आकारमान शहामृगाएवढे होते. त्यांचे पाय व मान लांब होती. डोके लहान व पुढील पाय अगदी आखूड व कमजोर होते. त्यांना दात नव्हते व त्यांना पक्ष्यांप्रमाणे चोच असावी. क्रिटेशसपूर्वीची त्यांची निश्चित माहिती उपलब्ध झालेली नाही.
सॉरोपोडा उपगणाचे प्राणी हे शाकाहारी चतुष्पाद सॉरिश्चिया असून त्यांचे पूर्वज बहुधा थेरोपोडांच्या ट्रायासिक कालीन प्लेटिओसॉरिडी कुलाचे प्राणी असावेत. दक्षिण आफ्रिकेतील यांच्या जीवाश्मांच्या ट्रायासिक व जुरासिक प्रारंभीच्या नोंदी अपूर्ण आहेत परंतु त्यांच्या उत्तर जुरासिक व पूर्व क्रिटेशस कालीन जीवाश्मांची अधिक चांगली माहिती इंग्लंड, उत्तर फ्रान्स व अमेरिकेच्या संयुक्त संस्थानांचा पश्चिम भाग येथून उपलब्ध झाली आहे. टांगानिका, मॅलॅगॅसी, भारत, चीन, ऑस्ट्रेलिया, ब्राझील व पॅटागोनिया येथे यांचे विपुल जीवाश्म आढळले आहे. या प्राण्यांचे दात लांब, दंडगोलाकार वा चमच्याच्या आकाराचे आणि धारदार कडांचे होते. ग्रैव (मानेतील) व पृष्ठीन मणके पश्चगर्ती होते. पायांची हाडे जवळजवळ भरीव होती, पुढचे पाय मागच्या पायांपेक्षा फारसे आखूड नव्हते. सर्व पायांना पाच बोटे होती. त्यांना त्वचीय चिलखत नव्हते.यातील प्राण्यांचे आकारमान सामान्यतः मोठे होते, काही तर राक्षसी आकारमानांचे होते. उदा.,डिप्लोडोकसच्या काही जाती २५-३० मी. लांब, ब्रॉंटोसॉरसच्या काही जाती २० मी.पेक्षा जास्त लांब, तर ब्रँकिओसॉरस सु.२८ मी. लांब होता. सॉरोपोडांचा प्रसार विस्तृत झालेला असल्याने ते जुरासिक व क्रिटेशस कालीन खडकांच्या थरांचे सहसंबंध ठरविण्यास उपयुक्त आहेत.
मध्य प्रदेशातील उत्तर क्रिटेशस कालीन खडकांत सॉरोपोडांचे अंटार्क्टोसॉरस, टिटॅनोसॉरस व लाप्लेटासॉरस या वंशांचे तर दक्षिण भारतातील अरियालूर मालेत टिटॅनोसॉरस वंशाचे जीवाश्म सापडले आहेत.
(आ) ऑर्निथिशिया: सॉरोपोडांप्रमाणे ऑर्निथिशिया गटातील डायनोसॉर शाकाहारी होते व सामान्यतः ते मोठ्या आकारमानाचे होते. तथापि हे प्राणी त्यांच्या राक्षसी आकारमानाऐवजी विविधतेसाठी जास्त प्रसिद्ध आहेत. त्यांचे दात चापट, चमच्याच्या आकाराचे व दातांच्या कडा सूक्ष्मदंती होत्या. पुष्कळांना पुढचे दात नसत परंतु त्यांना पक्ष्यांप्रमाणे शृंगी चोच असावी. मणके पश्चगर्ती, दोन्ही टोके सपाट असलेले किंवा कधीकधी दोन्ही बाजू अंतर्गोल असलेले होते. पाय भरीव किंवा कधीकधी पोकळ होते. कवटी जवळजवळ पूर्णपणे हाडांचा बनलेली असे. काहींमध्ये त्वचीय चिलखत चांगले विकसित झालेले होते असे चिलखत नसलेले प्राणी बहुतकरून द्विपाद होते. मात्र चिलखत असलेल्या प्राण्यांचे आधीचे वंश कदाचित द्विपाद व नंतरचे चतुष्पाद होते. या गटाचा काळ जुरासिक ते क्रिटेशस असा आहे.
ऑर्निथोपोडा उपगणातील बहुतेक प्राणी द्विपाद होते. त्यांना लांब व बळकट शेपटी होती व इतर ऑर्निथिशियांप्रमाणे यांची कवटी लहान नव्हती. त्वचीय चिलखत नव्हते अथवा अल्पविकसित होते. पूर्व जुरासिकमध्ये याचे थोडेच जीवाश्म आढळतात मात्र दोन्ही अमेरिका, दक्षिण आफ्रिका, टांगानिका, इंग्लंड, जर्मनी, ऑस्ट्रिया, फ्रान्स, हंगेरी, मंगोलिया इ. प्रदेशातील उत्तर जुरासिक व क्रिटेशस कालीन खडकांत या गटाचे पुष्कळ जीवाश्म आढळलेले आहेत. भारतामध्ये या उपगणापैकी ब्रँकिपोडोसॉरस वंशाचे जीवाश्म मध्य प्रदेशातील लॅमेटा थरांत सापडले आहेत.
ऑर्निथिशियांपैकी स्टेगोसेफॅलिया उपगणाचे प्राणी सर्वांत आधी अवतरले. स्टेगोसेफॅलिया चतुष्पाद होते आणि त्यांच्या पाठीवर व शेपटीवर संरक्षक पट्टांच्या व कंटकांच्या दोन ओळी होत्या. हे प्राणी मोठे होते व स्टेगोसॉरससारख्या काहींची लांबी सु.८ मी.होती. उत्तर अमेरिका, इंग्लंड, नॉर्मंडी (फ्रान्स), टांगानिका इ. प्रदेशांतील जुरासिक कालीन खडकांतच या उपगणाचे जीवाश्म सापडले आहेत. चतुष्पाद ऑर्निथिशियांपैकी अँकिलोसॉरिया या गटातील प्राणीही मोठ्या चणीचे होते. त्यांना भक्कम चिलखत होते व ते एखाद्या प्रचंड कासवांसारखे दिसत असत. त्यांची कवटी सापेक्षतः रुंद व मोठी होती. काहींना दात नव्हते. त्यांचे जीवाश्म मुख्यत्वे दोन्ही अमेरिका, दक्षिण आफ्रिका व इंग्लंड या भागांतील क्रिटेशस व विशेषतः उत्तर क्रिटेशस कालीन खडकांत आढळले आहेत. सेरॅटॉप्सिया गटाचे प्राणी मोठ्या चणीचे व चतुष्पाद होते. त्यांचे पुढील व मागील पाय जवळजवळ सारख्या लांबीचे होते. त्यांच्यापैकी काहींना, विशेषतः सेरॅटॉप्सिडी कुलातील प्राण्यांना अस्थिमय गाभा असलेली शिंगे होती. सेरॅटॉप्स व ट्रायसेरॅटॉप्स हे यांचे अधिक माहिती झालेले वंश आहेत.
(१४) टेरॉसॉरिया गण : निश्चितपणे टेरॉसॉरियाचे असलेले सर्वांत आधीचे जीवाश्म यूरोपातील पूर्व जुरासिक कालीन खडकांत सापडले आहेत. त्यांची पूर्वजपरंपरा निश्चितपणे माहित झालेली नाही परंतु स्कॉटलंडमधील उत्तर ट्रायसिक कालीन स्क्लेरोमॅक्लस हा छोटा चतुष्पाद डायनोसॉर ज्या वंशशाखेतील मानला जातो त्या वंशशाखेपासून बहुधा टेरॉसॉरिया निर्माण झाले असावेत. सर्वोत्कृष्टपणे टिकून राहिलेल्या त्यांच्या जीवाश्मांतही त्वचेवर पिसे वा खवले असल्याचे चिन्ह दिसत नाही. त्यांच्या सांगाड्यावरून त्यांच्यामध्ये जुरासिकच्या आधीच्या काळातही उड्डाणक्षमता आलेली दिसून येते व त्यांची हाडे या जीवनप्रणालीला योग्य अशी म्हणजे पोकळ होती मात्र त्यांच्या उड्डाणाचा पल्ला पक्ष्यांपेक्षा कमी होता. ज्यांना दात होते त्यांचे दात बारीक, तीक्ष्ण व अणकुचीदार असून अनियमितपणे मांडणी असलेले व पुढे वाकडे झालेले व जबड्यांच्या कडांशी खळग्यांत रोवलेले असत. अग्रत्रिकास्थीय मणके अग्रगर्ती व शेपटीतील मणके दोन्ही बाजूंनी अंतर्गोल होते. पुढच्या पायांचे पंखासारख्या संरचनेत परिवर्तन झालेले होते पुढच्या पायाचे फक्त पाचवे बोट अतिशय लांबट झालेले होते आणि पंखाच्या पटलाला आधार देण्यासाठी ते परिवर्तित झालेले होते. मागील पाय डायनोसॉरांप्रमाणे होते व मागील पायाच्या २ ते ४ बोटांना नख्या होत्या. पुढच्या पायांचे पंखांमध्ये परिवर्तन झाल्याने त्यांना मागील पायांवर चालणे बहुधा शक्य होत नसावे व त्यामुळे पुढील पायांच्या बोटांनी झाडांना टांगून घेण्याची सवय त्यांनी आत्मसात केली असावी. आधीच्या व उत्तर जुरासिकमधील काहींनाही लांब शेपटी होती पण क्रिटेशसमधील टोरोसॉराची शेपटी आखूड होती. यांची कवटी लहान, लांबोडकी व अग्र बाजूस पक्ष्यांप्रमाणे कमीअधिक निमुळती अथवा गोलसर टोक असलेली निमुळती अशी होती,तर पश्च बाजूला टेरॅनोडॉनमधल्याप्रमाणे ती चंद्रकोरीच्या आकाराची होती. मस्तिष्क-गुहा संरचनेच्या दृष्टीने पुष्कळशी पक्ष्यांसारखी होती पण ती कवटीच्या आकारमानाच्या मानाने अधिक लहान होती. पेलिकनप्रमाणे टोरोसॉरांना खालच्या जबड्याखाली अधोहनु-कोष्ठ होता आणि टोरोसॉर बहुधा कीटक व मासे खात असावेत.
टोरोसॉरियांचे जीवाश्म बहुतकरून सागरात व नदीमुखांच्या भागात साचलेल्या निक्षेपांत आढळतात. टोरोसॉर पूर्व जुरासिक काळापासून अस्तित्वात होते, क्रिटेशसमध्ये त्यांच्या विकासाने कळस गाठला व क्रिटेशसच्या शेवटी ते निर्वंश झाले. यूरोपच्या बाहेरील भागांत यांचे जुरासिक कालीन जीवाश्म विरळाच आढळतात. मात्र क्रिटेशसमध्ये त्यांचा विस्तृत प्रमाणात प्रसार झाला होता. सर्वांत चांगले टिकून राहिलेले क्रिटेशन कालीन काही जीवाश्म उत्तर अमेरिकेत आढळले आहेत. त्यांचे आकारमान चिमणीपासून ते राक्षसी पक्ष्यांएवढेही होते. काहींच्या पंखांचा विस्तार १.५ मी. व त्यापेक्षाही जास्त होता,उदा.,टोरॅनोडॉन ऑक्सिडेंटॅलिसच्या पंखांचा विस्तार ६ मी.होता. ऑर्निथोकीरिडी कुलामध्ये माहीत असलेले सर्वांत टोरोसॉर येतात. टोरॅनोडॉन ऑक्सिडेंटॅलिस हा उडणाऱ्या सरीसृपांपैकी माहीत असलेला सर्वंत मोठा सरीसृप आहे.
टेरोसॉर पुष्कळसे पक्ष्यांसारखे दिसत असले, तरी त्यांची काही वैशिष्ट्ये डायनोसॉरांप्रमाणे होती. उदा., त्यांचे मागचे पाय आणि कवटीचे काही घटक टेरोसॉर हे पक्ष्यांचे पूर्वज नाहीत.
टेरोसॉरियांचे दोन उपगण करतात : (१) टेरोडर्माटा (किंवा ऱ्हँपिऱ्हिंकॉइडिया) यांची शेपटी लांब होती. पूर्ण जबड्यांत नेहमी दात असत. हे सर्व सामान्यकृत व अदिम प्रकारचे टेरोसॉर होते. यांचे जीवाश्म जर्मनी, बव्हेरिया, इंग्लंड व पूर्व आफ्रिका या प्रदेशांतील जुरासिक कालीन खडकात आढळले आहेत. (२) ऑर्निथोकीरॉइडिया (किंवा टेरोडॉक्टिलॉइडिया) यांची शेपटी आखूड होती. जबड्याच्या फक्त अग्र भागात दात असत किंवा दात नसतही. यांचे जीवाश्म उत्तर जुरासिक व क्रिटेशन कालीन खडकांत आढळले आहेत.
बव्हेरियातील उत्तर जुरासिक कालीन लिथोग्राफिक खडकांत आढळलेल्या ॲनुरोग्नॅथसचा अपूर्णावस्थेत टिकून राहिलेला एकमेव सांगाडा हा संशयास्पद असा जीवाश्म आहे. पुष्कळ बाबतींत याचे टेरोडर्माटा उपगणाशी साम्य आहे मात्र याची शेपटी ऑर्निथोकीरॉइडियांप्रमाणे आखूड आहे.
पक्षी वर्ग : [⟶ पक्षि वर्ग]. बव्हेरियातील उत्तर जुरासिक कालीनलिथोग्राफिक खडकांत सापडलेले ⇨ आर्किऑप्टेरिक्स व आर्किऑर्निस हे पक्ष्यांचे सर्वांत जुने जीवाश्म आहेत, ते आकाराने कावळ्याएवढे असून त्यांच्याभोवती त्यांच्या पिसांचे ठसे उमटलेले आहेत. मात्र आधुनिक पक्ष्यांत नसलेली काही वैशिष्ट्ये त्यांच्यात होती. ती अशी त्यांना दात होते, त्यांची शेपटी शरीराच्या मानाने पुषकळच लांब होती जवळजवळ त्रिकास्थीपर्यंतचे दोन्ही बाजूंनी अंतर्गोल असलेले आदिम मणके मुक्त होते आणि त्यांची हाडे वायवीय म्हणजे पोकळ नव्हती. त्यांच्या सांगाड्याच्या वैशिष्ट्यांवरून ते वृक्षवासी व उड्डाणाचा पल्ला कमी असलेले पक्षी होते, असे दिसते. त्यांची पूर्वपरंपरा निश्चितपणे माहीत नसली, तरी ते निसंशयपणे लहान आर्कोसॉरिया सरीसृपांपासून आले असावेत व एकेकाळी समजण्यात येई त्याप्रमाणे ते उडणाऱ्या सरीसृपांच्या टेरोसॉरिया या वंशशाखेपासून आलेले नाहीत.
क्रिटेशसमध्ये सुद्धा पक्ष्यांच्या जीवाश्मांच्या नोंदी अतिशय अपूर्ण व विरळच अशा आहेत. यांपैकी कॅनझस (अमेरिका) येथील उत्तर क्रिटेशन कालीन नायाब्रेरा शैलसमूहातील इक्थिऑर्निस व हेस्परॉर्निस हे जीवाश्म अधिक चांगले माहित झाले आहेत. त्यांना दात होते. इक्थऑर्निसला बळकट कणी असलेली उरोस्थी (छातीचे हाड) होती. तीवरून तो उडणारा पक्षी असल्याचे सूचित होते. हेस्परॉर्निस हा पोहणारा होता व त्याची उरोस्थी कमजोर व कणाहीन होती. मात्र त्याचे पाय मजबूत होते आणि पाय घोट्यापासून बाजूला पसरता येऊ शकत होते व तशी स्थितीत त्यांचा पोहण्याकरिता वल्ह्यांप्रमाणे वापर करता येत असावा.
क्रिटेशन कालीन बहुतेक पक्ष्यांना दात होते. त्यांच्या सांगाड्यांची इतर वैशिष्ट्ये मात्र आधुनिक पक्ष्यांसारखी होती उदा., भुजास्थी, ऊर्वस्थी (मांडीचे हाड) व कवटीची हाडे वायवीय होती म्हणजे त्यांच्या हवायुक्त पोकळ्या व पातळ भित्ती होत्या.
नवजीव काळातील पक्ष्यांना दात नव्हते व त्यांच्या सांगड्याची हाडे वायवीय होती. त्यांच्यापैकी कित्येकांना उडता येत नव्हते. त्यांपैकी काहींची पूर्वपरंपरा बहुधा क्रिटेशस काळाइतकी मागे गेलेली आढळते. उडणाऱ्या पक्ष्यांच्या तुलनेने जलचर पक्ष्यांचे जीवाश्म अधिक चांगले टिकून राहिलेले आहेत कारण उडणाऱ्या पक्ष्यांचा शत्रूद्वारे व क्षयाद्वारे पुष्कळ प्रमाणात नाश होतो. तसेच असुरक्षित नैसर्गिक परिसरात त्यांचा जास्त नाश होतो व एकूनच पक्ष्यांचे जीवाश्म तुटपुंजे असल्याने स्तरवैज्ञानिक सहसंबंध ठरविण्यासाठी त्यांची अल्पशीच मदत होते. पक्षी हे उडत सतत स्थानांतरण करणारे असल्यामुळे त्यांच्यावरून पूर्वीच्या भूप्रदेशांतील दुवा आणि पूर्वीच्या काळातील प्क्ष्यांच्या प्रसाराची तऱ्हा यांचा मागोवा घेण्यास त्यांच्या जीवाश्मांचा उपयोग होऊ शकत नाही. तसेच त्यांच्या अपूर्णावस्थेत टिकून राहिलेल्या जीवाश्मांद्वारे त्यांच्यातील आप्तभाव शोधून काढणे पुष्कळदा अवघड असते. तथापि त्यांच्या जीवाश्मांवरून सर्वसामान्यपणे पूर्वी जलवायुमानात (दीर्घकालीन सरासरी हवामानात) झालेल्या बदलांची कल्पना येऊ शकते. क्रिटेशस काळातील पक्षी निश्चितपणे आदिम प्रकारचे होते, तथापि नवजीव काळातील पक्षी हे पुष्कळ प्रमाणात त्यांच्या आधुनिक नातेवाईकांसारखे होते. सांगाड्याच्या अवशेषांव्यतिरिक्त अंड्यांचे (किंवा त्यांच्या घाटांचे) जीवाश्मही क्रिटेशस व नवजीव काळांतील निक्षेपांत सापडले आहेत.
इतर भागांतल्याप्रमाणे भारतातील पक्ष्यांचे जीवाश्मही कसेबसे संपूर्णपणे टिकून राहिलेले आहेत. शिवाय ते तुटपुंजे आहेत व कधीकधी तर त्यांच्या वर्गीकरणातील योग्य स्थानाविषयी संदिग्धता आढळते. तथापि पंजाबातील प्लायोसीन कालीन उत्तर शिवालिक शैलसमूहांत ड्रोमीओग्नॅथी गणातील स्टुथिओ व ड्रोमीअस आणि यूऑर्निथिस गणातील पेलिकॅनस व लेप्टोप्टिलस हे जीवाश्म सापडले आहेत.
पक्ष्यांचे वर्गीकरण पुढीलप्रमाणे केले जाते पॅलीऑर्निथिस गटात जुरासिक कालीन प्रचीन पक्षी येतात. याचे पुढील आणखी तीन उपगट केले जातात (१) ओडोंटोग्नॅथी यात उत्तर क्रिटेशस काळातील इक्थिऑर्निस-हेस्परॉर्निस यांच्याशी संबंधित असलेला आदिम गट येतो, (२) पॅलिओग्नॅथी यात शहामृग, किवी इत्यादींसारखे न उडणारे पक्षीच येतात आणि (३) नीओग्नॅथी यात उडणारे पक्षी व त्यांच्यापासून अपकृष्ट (कमी विशेषीकृत) झालेले पक्षी येतात. पुष्कळदा यांच्या उरोस्थीवर कणा असतो. पॅलीओग्नॅथीमधील पुष्कळ पक्षी प्रचंड आकारमानाचे होते,उदा.,न्यूझीलंडमधील डायनॉर्निस पक्ष्याची उंची ४ मी. नीओग्नॅथीमधील फोरोऱ्हॅकॉस पक्ष्यांची उंची सु.२ मी. होती.
स्तनी वर्ग : [⟶ स्तनी वर्ग].सस्तन प्राणी नियततापी (ज्यांच्या शरीराचे तापमान परिसराच्या तापमानापेक्षा जास्त असून स्थिर असते असे) असून पृष्ठवंशीपैकी ते सर्वांत जास्त प्रगत प्राणी आहेत यांच्या पैकी मोनोट्रिमॅटा या अदिम छोट्या गटातील प्राणी अंडी घालणारे आहेत परंतु बहुसंख्य सस्तन प्राणी पूर्ण विकसित अशा पिल्लांना जन्म देतात. मार्सुपिएलिया या आदिम छोटया गटातील मादी मात्र काही काळपर्यंत आपल्या पिलाला शिशुधानीमध्ये (जिच्यात पिल्लाची वाढ पूर्ण होते अशा मादीच्या उदरावरील पिशवीमध्ये) ठेवून आपल्याबरोबर नेते. सर्व सस्तन प्राण्यांच्या पिलांची वाढ मातेच्या दुधावर होते. मासे, उभयचर व सरीसृप यांच्या उलट बहुतेक सस्तन प्राण्यांच्या शरीरावर खवले, गाठी, प्रशल्क इत्यादींचे आवरण नसते एडेंटाटा-गणातील काही प्राणी मात्र याला अपवाद आहेत. सस्तन प्राण्यांच्या सर्व अंगावर किंवा काही भागांवर इतर भागापेक्षा जास्त केस असतात अथवा सीटॅसिया (देवमासे व डॉल्फिन) व सायरेनिया (समुद्र-गायी) या गणांतील प्राण्यांप्रमाणे त्यांची त्वचा अनावृत असते.
सस्तन प्राण्यांचा सांगाडा पूर्णपणे हाडांचा बनलेला असतो. काही कवट्या व सांगाड्यांचे काही भाग हे चांगल्या प्रकारे टिकून राहिलेले असून त्यांच्यावरून कमीअधिक प्रमाणात सस्तन प्राण्यांच्या जीवाश्मांची फेरमांडणी करता येणे शक्य असते. तथापि दातांची झीज कमी प्रमाणात होत असल्यामुळे सस्तन प्राण्यांच्या अवशेषांमध्ये दात सर्वांत जास्त प्रमाणात आढळतात. तसेच त्यांच्या पायांची हाडे पूर्णतया अस्थिमय असल्याने अशी हाडे सामान्यतः ती जेथे सांधलेली असतात त्या पृष्ठांच्या वैशिष्ट्यांसह चांगली टिकून राहिलेली असून प्राणी ओळखण्याच्या दृष्टीने ती उपयुक्त असतात. यामुळे सस्तन प्राण्यांच्या जीवाश्मांच्या वर्गीकरणविषयक कामात दात व पायाची हाडे चांगली उपयुक्त ठरली आहेत.
सरीसृप व उभयचर यांच्या उलट सस्तन प्राण्यांचे चार पाय त्यांच्या शरीराच्या पातळीत व धडाखाली असतात. अशा प्रकारे पायांमुळे शरीराला अधिक चांगल्याप्रकारे आधार मिळतो आणि हालचालही अधिक चांगल्याप्रकारे करता येते.
सस्तन प्राणी मुख्यत्वे भूचर आहेत सीटॅसियांसारखे काही जलचर तर किरोप्टेरा (वटवाघळे) उडणारे आहेत. या प्राण्यांच्या पायांत त्यांच्या जीवनप्रणालीस योग्य असे परिवर्तन झालेले असते. सरीसृपांशी तुलना केल्यास सस्तन प्राण्यांच्या कवटीतील घटक थोडेच असतात कशेरुक-अक्षाला जोडणारे दोन अस्थिकंद (अस्थींवरील खळग्यात बसण्यायोग्य वाटोळे उंचवटे) कवटीला असून पश्चकपालाचे पृष्ठ उभट असते आणि पार्श्वकपाल व ललाटीय भाग वर उचलले गेलेले असतात. खालच्या जबड्यातील प्रत्येक बाजूला एकच दंतिकास्थी असून दात जबड्याच्या कडांशी असतात आणि खालचा जबडा कवटीशी सरळ सांधला गेलेला असतो हनुसंधिकेद्वारे जोडलेला नसतो.
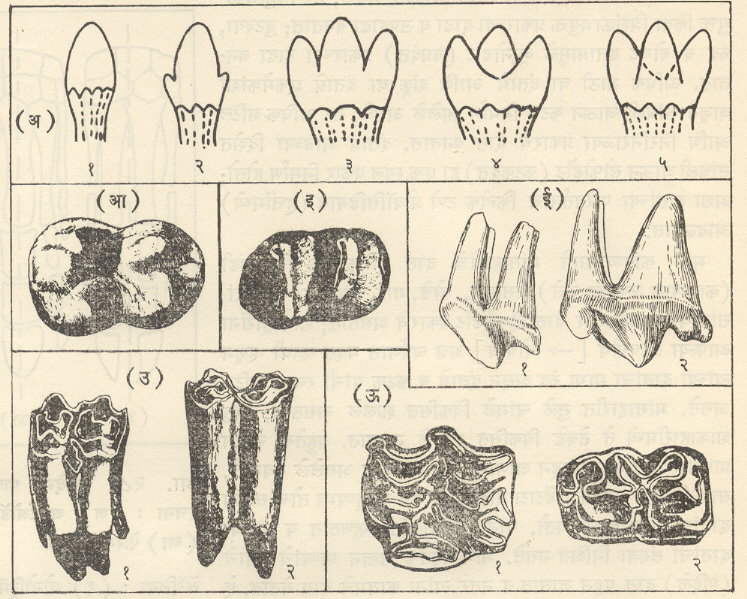
मणक्यांच्या दोन्ही बाजू सपाट असून लगतच्या दोन मणक्यांमध्ये तंतु-उपास्थिमय (जिचे आधारद्रव्य मुख्यत्वे तंतूचे बनलेले असते अशा कूर्चेची) चकती असते. तथापि काही खुरी प्राण्यांमधील मानेचे मणके पश्चगर्ती असतात. सस्तन प्राण्यांच्या पृष्ठवंशाचे पाच भाग पाडता येतात या प्रत्येक भागातील मणक्यांची संख्या सामान्यतः कमी जास्त पण थोडीच असते. अशाच प्रकारे मानेच्या भागात सामान्यपणे ७ मणके असतात वक्षाच्या भागात १२ ते १४, कटी भागात ५ ते ७ आणि त्रिकास्थीय (पृष्ठवंशाच्या टोकाच्या हाडाच्या) भागात २ ते ५ मणके असतात. शेपटीच्या भागात अनेक मणके असतात परंतु त्यांची संख्या कमी होत जाण्याची प्रवृत्ती आढळते. सीटॅसिया व सायरेनिया यांच्यात त्रिकास्थीय भाग वेगळा मानला जात नाही.
बहुतेक सस्तन प्राण्यांमध्ये दाताच्या गोर्द-गुहेभोवती असलेल्या दंतिनाच्या थराभोवती दंतवल्काचा थर असतो. माथ्यावरील दंतशीर्ष व कटक यांच्यातील खोलगट भाग संधानकाच्या (सिमेंटच्या) कमीअधिक जाड थराने भरलेला असतो आणि दंतमूलांवर संधानकाचा पातळ थर असतो. बुटक्या माथ्याच्या दातांना ब्रॅकिओडोंट (लघुदंत) तर गोर्द-गुहा मोकळी असलेल्या दंडगोलाकार अथवा प्रचिनाकार दातांना हिप्सोडोंट (तुंगदंत) म्हणतात.
सस्तन प्राण्यांच्या दातांचे कृंतक, सुळे, उपदाढा व दाढा असे वेगळे प्रकार करतात. अशा विभेदित दातांची प्रवृत्ती थेरोमॉर्फा सरीसृपांच्या सायनोडोंशिया या उपकुलातील काही प्राण्यांतही आढळते. मोनोट्रिमॅटाच्या आदिम गटांतच सरीसृपांसारखे अविभेदित शंक्वाकार दात आढळतात. दातांचे असे विभेदन दातांना कराव्या लागणाऱ्या कार्याशी (उदा., भक्ष्य पकडणे, अन्न कापणे, त्याचे दलन करणे व भरडणे) निगडित आहे आणि ही विविध कार्ये तोंडात जेथे केली जातात तेथे त्या त्या प्रकारचे दात असतात.
हॅप्लोडोंट या एकच शंकू असलेल्या आदिम दातांपासून दोन्ही बाजूंना छोटे शंकू असलेले प्रोटोडोंट प्रकारचे दात आले आहेत. हे छोटे शंकू एकाआड एक वा सलग ओळीत असले, तर त्रिगुलिकायुक्त किंवा त्रिशंक्वाभयुक्त प्रकारच्या दाढा व उपदाढा बनतात बुटक्या, रुंद व बोथट दंताग्रामुळे ब्युनोडोंट (वप्रदंत) प्रकारच्या दाढा बनतात. अधिक गाठी वा दंताग्रे आणि शंकू वा दंताग्रे एकमेकांशी बाजूने सांधली जाऊन कटक निर्माण झालेले असले, तर अधिक जटिल आणि निरनिराळ्या प्रकारचे दात बनतात. दंताग्रे आडव्या दिशेत सांधली जाऊन लोफोडोंट (कटकदंत) हा एक स्थूल प्रकार निर्माण होतो अशा दातांच्या परिवर्तनाचे कित्येक टप्पे प्रोबॉसिडियात (हत्तींमध्ये) आढळतात.
मांस कापण्यासाठी मांसाहारींचे दात गुलिकायुक्त त्रिज्यखंडी (कापण्यास उपयुक्त असे) असतात. घोडे, गाई, म्हशी इ. जनावरांसारख्या शाकाहारींचे दात हिप्सोडोंट प्रकारचे असतात शाकाहारींना आपल्या ओषधीय [⟶ ओषधि] अन्न चर्वणास मदत व्हावी म्हणून त्यांच्या दातांचा माथा रुंद असून दंताग्रे व कटक यांची रचना जटिल असते. मांसाहारींत सुळे चांगले विकसित झालेले असतात परंतु शाकाहारींमध्ये ते तेवढे विकसित झालेले नसतात. बहुतेक सस्तन प्राण्यांचे दात एकदा पडून जाणारे असतात. दात असलेले देवमासे, सायरेनिया व बहुतेक एडेंटाटा यांच्यामध्ये सर्व आयुष्यभर तोच साध्या दातांचा संच कामी येतो, यांना एकवारदंती म्हणतात व त्यांच्या दातांची संख्या निश्चित नसते. मात्र बहुतेक सस्तन प्राण्यांचे दुधाचे (पहिले) दात पडून जातात व नंतर त्यांना कायमचे दात येतात, हे द्विवारदंती असून त्यांच्यातील प्रत्येक गटातील प्राण्याच्या दातांची संख्या ठराविक असते. द्विवारदंतीपैकी आदिम प्राण्यांना ४४ पर्यंत कायमचे दात होते. तथापि प्रत्येक गणात व कुलात दातांची संख्या कमी होत जाण्याची प्रवृत्ती आढळते काहींमध्ये तर दात नसणे हेच वैशिष्ट्य ठरते.
प्रत्येक जबड्यातील दातांची मांडणी सममित (सारखी) झालेली असते मात्र दोन्ही जबड्यांत वेगवेगळी असू शकते. एकाच बाजूच्या प्रत्येक प्रकारच्या दाताचा आकडा देऊन सस्तन प्राण्यांचा दंतविन्यास देण्याची प्रथा आहे आणि एकूण दात हे सूत्रात दिलेल्या दातांच्या दुप्पट असतील. अशा प्रकारे मानवाच्या बाबतीत हे सूत्र पुढीलप्रमाणे देतात
| वरचा जबडा | = | कृंतक २, | सुळा १, | उपदाढा २, | दाढा ३ | = |
| खालचा जबडा | कृंतक २ | सुळा १, | उपदाढा २, | दाढा ३ |
८/८× २ म्हणजे ३२. हे सूत्र संक्षिप्तपणे असे लिहितात २.१.२.३ =
२.१.२.३
८/८× २ म्हणजे१६/१६ म्हणजे ३२. हरिणाच्या बाबतीत हे सूत्र
०.१.३.३/३.१.३.३ = ७/१२ × म्हणजे १४/२० म्हणजे ३४ असे आहे.
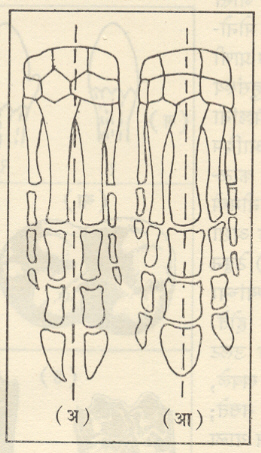
सस्तन प्राण्यांच्या क्रमविकासातील दुसरे महत्त्वाचे वैशिष्टय म्हणजे आदिम प्रकारच्या पंचांगुली (पाच बोटे असलेल्या) पायाची बाजूची बोटे क्रमाने कमी होत जाणे. जीवनप्रणालीशी जळवून घेण्यासाठी हे अनुकूलन घडून आले व काही नको असलेले घटक नंतर वापरले न गेल्याने त्यांचा ऱ्हास होऊन नाहीसे होत गेले. मांसाहारींत ४ किंवा कधीकधी ५ बोटे असतात, तर नोटोअंग्युलेटात ३ पर्यंतही बोटे असतात. खुरी (अंग्युलेटा) प्राण्यांच्या समखुरी (पेरिसोडॅक्टिला) या शाखेत शेवटी जनावरांप्रमाणे दोन खुरी तर विषमखुरी (आर्टिओडॅक्टिला) प्राण्यांत घोड्याप्रमाणे एक खुरी प्राण्यांचा टप्पा येतो [⟶ अंग्युलेटा]. भक्ष्य पकडण्यासाठी मांसाहारींना नख्या आल्या. खुरी प्राणी सामान्यतः ज्या कठीण पृष्ठभागावरून हिंडत त्यावर चालणे सोयीचे व्हावे म्हणून त्यांना खूर आले. गाई, म्हशी इत्यादींसारख्या प्राण्यांना चिकट चिखलातून चा लण्याच्या दृष्टीने दोन खूर सोयीचे होते. सीटॅसिया, पिन्निपीडिया (सील व वॉलरस) व सायरेनिया यांच्या पुढील पायांचे त्यांच्या जलचर जीवनप्रणालीला अनुरुप अशा वल्ह्या सारख्या रूपात परिवर्तन झालेले आहे. ( मागील पायांचा कमीअधिक प्रमाणात क्षय झाला आहे) तर किरोप्टेरा प्राण्यांचे पुढील पाय पंखासारख्या संरचनेत परिवर्तित झालेले आहेत.
सस्तन प्राण्यांचे बहुतेक प्रमुख गट आधुनिक काळात राहत असून प्राणिशास्त्रज्ञांनी स्वीकारलेले त्यांचे वर्गीकरण भ्रूणविज्ञानावर आधारलेले आहे. त्यानुसार पुढीलप्रमाणे दोन उपवर्ग व त्यांचे गण केले जातात : (अ) इप्लॅसेंटॅलिया : (१) मोनोट्रिमॅटा (अथवा प्रोटोथेरिया), (२) मार्सुपिएलिया (किंवा मेटॅथेरिया) व (आ) प्लॅसेंटॅलिया (किंवा यूथेरिया) : याचे कित्येक गण आहेत. कित्येक गण, कुले इत्यादींमध्ये असलेले संबंध हे पूर्वी मानण्यात येत असल्यापेक्षा जीवाश्मांवरून अधिक जवळचे असल्याचे दिसून आल्याने वर्गीकरणावर जीवाश्मांपासून मिळालेल्या माहितीचा बराच प्रभाव पडला आहे. शिवाय मुख्यत्वे काही गटांच्या इतर गटांच्या संदर्भातील महत्त्वाच्या बाबतीत तज्ञांम ध्ये मतभेद आहेत व अशा प्रकारे काही उपगण हे गणाच्या दर्जाचे असल्याचे त्यांचे मत आहे. उदा., ॲग्युलेटा गणातील उपगण. तसेच काहींच्या मते सस्तन प्राणिवर्गाचे पुढील तीन उपवर्ग केले पाहिजेत . (१) मोनोट्रिमॅटा, मार्सुपिएलिया आणि (३) प्लॅसेंटॅलिया. मात्र या मतभेदांचा सस्तन प्राण्यांचे विविध गट वेगळे ओळखण्यावर क्वचितच परिणाम होतो.
इप्लॅसेंटॅलिया उपवर्ग : (१) मोनोट्रिमॅटा गण (अंडजस्तनी) : निश्चितपणे ओळखू येणारे मोनोट्रिमॅटा गणाचे जीवाश्म ऑस्ट्रेलियातील प्लाइस्टोसीन कालीन निक्षेपात आढळत असून ते हल्लीच्या पुढील वंशांपैकी असल्याचे म्हणता येतील : ऑर्निथोऱ्हिकस, एकिडूना व प्रोएकिडूना. अशा तऱ्हेने या गणाचे पुराप्राणिवैज्ञानिक महत्त्व थोडेच आहे. अंडी घालण्याची प्रवृत्ती व आदिम प्रकारचे शंक्वाकार दात ही सरीसृपांना जवळची असलेली वैशिष्ट्ये यांच्यात टिकून राहिलेली आहेत. आर्. ब्रूम व इतरांनी असे दाखवून दिले आहे की, थेरिओडोंट सरीसृपांच्या टायासिक कालीन इक्टिडिओसॉरिया या लहान आकरमानाच्या प्राण्याच्या गटातील प्राण्यांचे कवटीचे व सांगाड्याचे घटक मोनोट्रिमॅटांच्या अशाच घटकांपासून वेगळे ओळखणे अवघड आहे. अशा प्रकारे मोनोट्रिमॅटांचे इक्टिडिओसॉरियांशी अतिशय निकटचे संबंध असून बहुतकरून ते मोनोट्रिमॅटांचे पूर्वज असावेत. यामुळे जरी मोनोट्रिमॅटांचे जीवाश्म अपूर्ण व थोडेच असले, तरी पुराप्राणिवैज्ञानिकांना आणि प्राणिवैज्ञानिकांना त्यांच्याविषयी पुष्कळ औत्सुक्य आहे.
(२) मार्सुपिएलिया गण : [शिशुधान स्तनी ⟶ शिशुधानी]. हा इप्लॅसेंटॅलियांचा एक मोठा गट असून यामध्ये पुष्कळ विविधता आढळते. शिशुधान पिशावीला आधार देणारी शिशुधान-अस्थी व खालच्या जबड्याचा वाकलेला कोपरा ही या गणाची ठळक संरचनात्मक वैशिष्ट्ये होत यांमुळे ते इतर सस्तन प्राण्यांपासून वेगळे ओळखू येतात. सामान्यतः त्याचे पुढील पाय मागील पायांपेक्षा आखूड असतात. त्यांचे दात मांसाहारी, कीटकभक्षी किंवा शाकाहारी प्राण्यांसारखे असतात. बहुसंख्य शिशुधानींना ४ ते ६ दाढा असतात, कृंतक दात पुष्कळदा कृंतक (कुरतडणाऱ्या) प्राण्यांसारखे [⟶ कृंतक गण] असतात व प्रत्येक जबडयात एक कृंतक दात विशेष वाढलेला असतो सुळे बळकट असून बहुधा त्यांना दोन दंतमूले असतात. त्यांच्या मागील पायांना ४ वा ५ बोटे असतात आणि त्यांची संख्या कमी होण्याची प्रवृत्ती आढळते. या प्राण्यांचे पूर्वज पाच बोटांचे वृक्षवासी प्राणी असावेत, असे मानतात.
शिशुधानींचे सर्वांत पुरातन जीवाश्म यूरोप व दक्षिण आफ्रिका या भागांतील उत्तर जुरासिक कालीन खडकांत सापडले आहेत. दोन्ही अमेरिका व यूरोप यांच्यातील जुरासिक, क्रिटेशस व तृतीय कालीन खडकांत यांचे जीवाश्म विस्तृतपणे पसरलेले असून ते विविध प्रकारचे आहेत. हल्लीच्या शिशुधानींपैकी डायडेल्फिडी (ऑपॉस्सम) कुलातील डायडेल्फिस व किरोनेक्टिस या वंशांचा प्रसार पॅटागोनिया ते कॅनडापर्यंतच्या भागात झाला आहे. सीनेलेस्टिडी कुल केवळ ऑस्ट्रेलियात आढळते मात्र यातील सीनोलेस्टीस वंश दक्षिण आफ्रीकेत आढळतो. हल्लीच्या कांगारूंचे फॅलॅंजेरिडी कुल तृतीय काळात ऑस्ट्रेलियात अवतरले व तेथेच राहिले. हल्लीचे इतर शिशुधानी फक्त ऑस्ट्रेलिया व त्यालगतच्या बेटांवरच आढळतात आणि तेथील प्लाइस्टोसीन कालीन निक्षेपांत त्यांचे जीवाश्म सापडतात.
ड्रोमॅथेरिआयडी कुल : (पॉलिप्रोटोडोंशिया उपगण) या लहान शिशुधानींचे अल्पसेच जीवाश्म आढळत असले. तरी ते शिशुधानी व ट्रायासिक कालीन सायनोडोंशिया सरीसृप यांच्यामधील दुवा दर्शविणारे पुरावा ठरले आहेत. पँटोथेरिआयडी कुलाचा (पॉलिप्रोटो डोंशिया उपगण) काळ जुरासिक ते उत्तर क्रिटेशस असा असून त्यांच्यापासून अपरास्तनींपैकी (प्लॅसेंटल वार असणाऱ्या सस्तन प्राण्यांपैकी) क्रिओडोंशिया (क्रव्यदंत) व इन्सेक्टिव्होरा (कीटकभक्षी) या गणांचे प्राणी निर्माण झाले असावेत.
प्लॅसेंटलिया उपवर्ग : (१) इन्सेक्टिव्होरा गण : अपरास्तनींपैकी हे सर्वांत आदिम प्राणी होत. सामान्यतः ते लहान, निशाचर, नमुनेदार भूचर, पुष्कळ वेळा बिळात राहणारे व काही वृक्षवासी आहेत. त्यांचे डोके कमी दबलेले (खोलगट) असून पुढचे पाय पादतलचारी (पूर्ण तळपाय जमिनीली टेकून चालणारे) व बहुतकरून पाच बोटे असलेले असतात. बिळे करणाऱ्या प्राण्यांच्या पुढील पायांचा खणण्याच्या दृष्टीने अनुरूप असा खास विकास झालेला असतो. दातांचे सर्वसामान्य सूत्र ३.१.४.३/३.१.४.३ असून पुष्कळदा सुळे व उपदाढा वेगळ्या ओळखणे अवघड असते. वरच्या दाढा सामान्यपणे त्रिगुलिकायुक्त असून त्यांना जोडणारे कटक असतात.
या गणाचे शिशुधानींच्या पँटोथेरिआयडी कुलाशी दूरचे नाते असल्याचे मानतात. हल्लीचे कीटकभक्षी केवळ पूर्व गोलार्धात व उत्तर अमेरिकेत आढळतात. त्यांचे जीवाश्मही त्याच प्रदेशांत आढळतात. श्रोणिप्रतरसंधीच्या स्वरुपांनुसार केलेल्या दोन गटांपैकी मेनोटायफ्ला गटाचे ट्युपेइडी कुल पूर्व आशियात व मॅक्रोस्केलिडी कुल आफ्रिकेत आढळत असून त्यांच्या जीवाश्मांच्या नोंदी अनिश्चित आहेत. तथापि लायपोटायफ्ला या दुसऱ्या गटाच्या निर्वंश झालेल्या पुष्कळ प्राण्यांचे जीवाश्म यूरोपातील व उ. अमेरिकेतील पूर्व तृतीय कालीन खडकांत सापडले आहेत. कीटकभक्षींपैकी पुष्कळांचे नरवानरांशी (कदाचित हिप्सोडोंटिडी किंवा मिक्सोडोंटिडी कुलद्वारे), क्रियोडोंशियांशी व रोडेंशियांशी (कृंतक प्राण्यांशी प्लेसिओडॅपिडी कुलाद्वारे) नातेसंबंध असल्याचे दिसते.
(२) किरोप्टेरा गण : (वटवाघळे). हे अपरास्तनी आदिम प्रकारचे असले, तरी त्यांनी उडण्याची खास जीवनप्रणाली स्वीकारली आहे. त्यांचे पुढील पाय मागील पायांपेक्षा पुष्कळच लांब असून प्रबाहू लांब होऊन त्यांचे पंख्यासारख्या रचनेत परिवर्तन झालेले आहे तसेच पुढील पायांची बोटेही पुष्कळच लांबट झाली असून ती त्वचेने जोडली जाऊन पंख तयार झाले आहेत. त्यांचे मागील पाय व कटी अतिशय कमजोर असतात कृंतक दात आखूड झालेले असतात, पण सुळे नेहमी बळकट असतात वटवाघळे अधिक प्रमाणात कीटकभक्षी असल्याने या त्यांच्या प्रवृत्तीला जुळतील अशा मूलतः त्रिगुलिकायुक्त दाढा व उपदाढा त्यांना असतात.
यांच्या दोन गटांपैकी मेगॅकिरोप्टेरा हा गट लहान असून यातील प्राणी आकारमानाने मोठे व फलभक्षी (फळ खाणारे) आहेत. मायक्रोकिरोप्टेरा या दुसऱ्या गटातील प्राणी लहान व कीटकभक्षी आहेत. इओसीन काळाइतके पुरातन यांचे जीवाश्म आढळतात परंतु ऑलिगोसीन व मायोसीन काळातील जीवाश्मही थोडेच आढळतात. प्लाइस्टोसीन काळातील जीवाश्म मुख्यत्वे मायक्रोकिरोप्टेरा गटाचे असून ते हल्ली अस्तित्वात असलेल्या वंशाचे म्हणता येतील. ऱ्हेनोलोफस, व्हेस्पर्टिलियो इत्यादींसारखे हल्लीचे वंश मायोसीन काळापासून अस्तित्वात आहेत.
(३) कार्निव्होरा गण : (मांसाहारी). पिन्निपीडिया हा छोटासा जलचर प्राण्यांचा उपगण वगळता या गणातील प्राणी भूचर आहेत. त्यांचे आकारमान खोकडापासून वाघ-सिंहाएवढे असते. दात द्विवारदंती असून त्यांच्या दातांचे अधिक सर्वसामान्यपणे आढळणारे सूत्र ३.१.४.३/३.१.४.३ असे असते. दाढा व उपदाढा मूलतः त्रिगुलिकायुक्त असून सुळे चांगले बळकट व उलट वळलेले असतात. मांसाहारींचे पुढील तीन उपगण आहेत.
(क) क्रिओडोंशिया उपगण : (क्रव्यदंत). मांसाहारींपैकी हे सर्वांत आदिम प्रकारचे व सर्वांत आधी अवतरलेले प्राणी आहेत. त्यांची कवटी दबलेली व सामान्यपणे इतर शरीराच्या मानाने मोठी असते. खालच्या जबड्याच्या पश्च कोपऱ्याचा बाक हे शिशुधानींचे वैशिष्टय मेसोनायकिडी कुलातील काही प्राण्यांत आढळते. यावरून शिशुधानी हे क्रिओडोंशियाचे पूर्वज असावेत, असे सूचित होते. त्यांच्या पायांना चाराहून कमी बोटे नसतात. कधीकधी पाचवे बोट असते बोटांना खुरांसारख्या दोन बोथट नख्या असतात. ते बहुतकरून अंगुलिचारी (बोटांवर चालणारे) असून काही गट अर्धपादतलचारी (पायाच्या तळव्याचा काही भाग जमिनीवर टेकवून चालणारे) आहेत. इतर मांसाहारींप्रमाणे यांचे सुळेही बळकट व एक दंतमूल असलेले आहेत इतर दातांच्या संख्येत घट होण्याची प्रवृत्ती आढळत असली, तरी दातांचे सर्व प्रकार असतात. आर्क्टोसायॉनिडी या आदिम कुलात दाढा गुलिकासदृश-त्रिज्यखंडी असतात मेसोनायकिडी कुलात चुरा करणारे दात चांगले वाढलेले होते, मात्र ते नमुनेदार दारक (कापणाऱ्या) प्रकारचे नव्हते. भूचर मांसाहारींपैकी आकारमानाने खूपच मोठे प्राणी या कुलात जन्मले. हे कुल ऑलिगोसीन काळात अस्तित्वात होते. ऑक्सिईनिडी व हायईनोडोंटिडी कुलांमध्ये नमुनेदार दारक दात होते. मायसिडि कुलातील प्राणी लहान असले, तरी त्यांच्यातील मस्तिक-गुहा ही क्रिओडोंशियांमध्ये सर्वांत मोठी होती. या कुलाचे थोडेच जीवाश्म आढळतात. पॅलिओसीनच्या आधीपासून ते ऑलिगोसीनच्या सुरुवातीच्या काळात या कुलापासून फिसिपोडिया निर्माण झाले असावेत. पॅलिओसीनच्या आधीच्या काळापासून ते जवळजवळ पूर्ण ऑलिगोसीन काळात क्रिओडोंशियांचा विस्तृत प्रसार झाला होता. ऑलिगोसीनच्या अखेरीस फिसिपोडियांनी त्यांच्यावर जलदपणे मात केली पण ते भारतात व आफ्रिकेत मायोसीनमध्ये काही काळ टिकून राहिले होते.
(ख) फिसिपोडिया उपगण : या मांसाहारींचे मुस्कट काहीसे लांबट असून आदिम प्राण्यांत कवटी दबलेली पण अधिक प्रगत प्राण्यात ती फुगीर होत गेलेली आढळते. मस्तिष्क-गुहा मोठी व पीळ पडलेली असते. सुळे लांब, उलट वळलेले व बळकट असतात. दारक दात सामान्यपणे इतर दाढा व उपदाढांपेक्षा मोठे व लांब असतात त्यांना वरच्या जबड्यात तीन व खालच्या जबड्यात दोन दंतमूले असतात. बहुतेक फिसिपेडियांना चार बोटे असतात. फेलिडी व व्हायव्हेरिडी या कुलांतील प्राण्यांच्या नख्या प्रतिकर्षी (आत ओढून घेता येतील अशा) असतात.
या गणाची सर्व कुले हल्ली हयात आहेत व ती सर्व उत्तर इओसीनमध्येही अस्तित्वात होती काही ऑलिगोसीनमध्ये अवतरली.
कुत्रे, लांडगे, कोल्हे व खोकाड हे प्राणी कॅनिडी कुलातील असून हे कुल युरोपात उत्तर इओसीन काळात क्रिओडोंशियातील मायसिडी कुलापासून निर्माण झाले. उत्तर अमेरिकेत मध्य तृतीय काळात या कुलाचा कमाल प्रसार झाला होता तेथून हे प्राणी सर्व जगभर गेले व आता ते सार्वत्रिक झाले आहेत. विशेषतः कॅनिस वंशाचा प्रसार विस्तृत प्रमाणावर झाला असून त्यात पुष्कळ विविधता आढळते.
मांजरे, वाघ, सिंह बिबळ्ये व चित्ते हे प्राणी फेलिडी कुलातील असून ते अतिशय चपळ आहेत. या कुलाची पूर्वजपरंपरा अनिश्चित आहे परंतु हे कुल इओसीनच्या अखेरच्या किंवा ऑलिगोसीनच्या सुरूवातीच्या काळातील असले पाहिजे, कारण या कुलाचे म्हणता येतील असे जीवाश्म ऑलिगोसीन कालीन खडकांत सापडले आहेत. मांसाहारींमध्ये या कुलातील प्राण्यांचा नासा-प्रदेश (नाकाचा ओलसर शेंडा) लक्षात येईल इतक्याच अपुऱ्या लांबीचा असतो व कवटी उंच असते. यांचे दारक दात अतिशय कार्यक्षम असले, तरी कुलांच्या तुलनेने दाढांची संख्या कमी होण्याची प्रवृत्ती या कुलात अधिक आढळते. या कुलांमध्ये व विशेषतः यातील फेलिस वंशामध्ये अतिशय विविधता आढळते तसेच ऑस्ट्रेलिया व न्यूझीलंड वगळता सर्व जगभर याचा प्रसार झालेला आहे.
मुस्टेलिडी आणि व्हायव्हेरिडी (कस्तुरी मांजर) या कुलांतील प्राणी लहान पण अतिशय चपळ असून उत्तर गोलार्धात, विशेषतः दक्षिण यूरोप, आफ्रिका व आशिया या प्रदेशांत, त्यांचा विस्तृत प्रसार झालेला आहे.
अर्सिडी (अस्वल) हे कुल इतर फिसिपेडियांपेक्षा काहीसे वेगळे आहे. यातील प्राणी पादतलचारी, पाच बोटे असलेले व झाडावर चढण्यास सोयीच्या अशा नख्या असलेले आहेत. या कुलातील सर्वभक्षी व फलभक्षी प्राण्यांच्या उपदाढा दारक दातांपेक्षा मोठ्या असतात. हे कुल मध्य तृतीय काळात यूरोपमध्ये कॅनिडीतील सायनोडोंटिनी उपकुलापासून निर्माण झाले. मायोसीन व प्लायोसीन काळांत यातील प्राणी उत्तर अमेरिका व इतर प्रदेशांत गेले.
आधुनिक तरस हायनिडी कुलातील असून त्यांचे दात वैशिष्ट्यपूर्ण आहेत त्यांचे दारक दात बळकट असतातच, शिवाय दाढा लांब व त्यांचे शंक्वाकार टोक मजबूत असून त्यांचा हाडे फोडण्यासाठी उपयोग होतो. हे कुल बहुधा प्लोयोसीन काळात पिकेर्मी (ग्रीस) भागात अवतरले आणि नंतर त्याचा यूरोपात व आशियात प्रसार झाला. यातील बहुतेक प्राणी आता उत्तर गोलार्धात आहेत.
प्रोसिऑनिडी (रॅकून) कुलाचे प्राणी अमेरिकेतच आढळतात मात्र यातील आयल्युरस वंशाचे प्राणी हिमालयाच्या भागात अलगपणे राहत आहेत. याचे बहुतेक जीवाश्म यूरोप व दोन्ही अमेरिका या भागांतील मायोसीनपासूनच्या काळातील खडकांत आढळतात. मायसिडी कुलातील किंवा सायनोडोंटीनी उपकुलातील प्राणी या कुलाचे वंशज असावेत.
(ग) पिन्निपीडिया उपगण : यामध्ये सील व वॉलरस हे प्राणी येतात. या मांसाहारींचे पाय त्यांच्या जलचर जीवनप्रणालीस अनुसरून परिवर्तित झालेले आहेत पायाचे समीपस्थ घटक आखूड तर दूरस्थ घटक लांब व जाळीदार झालेले आहेत. उपदाढा व दाढा साध्या म्हणजे एकाच शंकूच्या अथवा उंच शंकू व त्यांच्या अग्र व पश्च बाजूस गौण दंताग्रे असलेल्या अशा असतात. कृंतक दात तीनपेक्षा कमी असतात पण सुळे चांगले वाढलेले असतात. या कुलाच्या उत्पत्तीविषयी निश्चित माहिती नाही परंतु मेंदूच्या संरचनेतील विशिष्ट साम्यांमुळे या कुलाचे अर्सिडी कुलाशी जवळचे नातेसंबंध असावेत, असे मानतात.
या कुलातील हल्लीचे प्राणी दोन्ही ध्रुवांभोवतीच्या प्रदेशात आढळतात. मायोसीन काळापासूनचे त्यांचे जीवाश्म अमेरिकेची संयुक्त संस्थाने (ऑरेगन व मेरीलँड), ईजिप्त, हंगेरी, ऑस्ट्रिया व दक्षिण रशिया या भागांत सापडले आहेत. ते यूरोपीय देश, दोन्ही अमेरिका न्यूझीलंड या भागांतील तृतीय काळाअखेरच्या आणि प्लाइस्टोसीन कालीन खडकांत आढळतात.
सीटॅसिया गण : देवमासे व शिंशुक (पॉरपॉइज) हे या गणातील प्राणी असून यातील प्राण्यांची हाडे सर्वसाधारण सच्छिद्र व स्पंजासारख्या पोताची असतात. कवटीची हाडे विशेष मोठ्या कोशिकांची (पेशींची) बनलेली असल्याने पातळ व हलकी असतात व ती जलचर जीवनप्रणालीस अनुरुप अशी असतात. कवटी धडापासून वेगळी नसते. नासा-प्रदेश लांब व अरूंद असून मस्तिष्क-गुहा उंचावली गेल्याने पश्च भागात तो उंच कटकासारखा दिसतो. कानाची हाडे (मध्यकर्ण व परिकर्ण) अतिशय भरीव असून ती इतर हाडांना सैलसरपणे जोडलेली असतात आणि म्हणून जीवाश्मरूपातील अवशेषांत पुष्कळदा सुटी आढळतात. मणक्यांच्या बाजू सपाट असून दोन मणक्यांच्या मध्ये उपास्थीचा जाड थर असतो. मानेत सामान्यतः ७ मणके असतात ते चपटे असून बॅलीनिडी (मिस्टॅकोसीटाय उपगण), डेल्फिनिडी (डॉल्फिन) व फायझिटेरिडी (ओडाँटोसीटाय उपगण) या कुलांतील काही प्राण्यांत मणके कमीअधिक प्रमाणात सांधले गेलेले असतात. वक्षामध्ये ९ ते १६, कटीत ३ ते २४ आणि शेपटीच्या भागात १८ ते ३० मणके असतात त्रिकास्थी नसते. इतर सस्तन प्राण्यांच्या उलट सीटॅसिया गणातील प्राण्यांमध्ये मणके आणि उरोस्थी यांना बरगड्या सैलपणे जोडलेल्या असतात. मिस्टॅकोसीटाय उपगणात उरोस्थी म्हणजे एक रुंद ढालीसारखी पट्टी असते परंतु आर्किओसीटाय व ओडाँटोसीटाय या इतर उपगणांत उरोस्थी दोन किंवा अधिक तुकड्यांची बनलेली असते.
यांना अनेक दात असून एकच दंतमूल असते बहुधा ते शंक्वाकार असून झ्युग्लोडोंटिडी (आर्किओसीटाय उपगण) आणि स्क्वॅलोडोंटिडी (ओडाँटोसीटाय उपगण) कुलांत मागच्या दातांना २ किंवा ३ दंतमूले असतात. पुढून मागील दिशेत दात नाहीसे होत जाण्याची प्रवृत्ती आढळते. जलचर जीवनप्रणालीत पर (फ्लिपर) म्हणून उपयोगी पडावेत या दृष्टीने पुढील पायांमध्ये परिवर्तन झालेले असते याकरिता भुजास्थी आखूड व बळकट झालेली असून दूरस्थ घटक लांब
होऊन पटलाने जोडलेले गेलेले असतात. स्कंधास्थी (खांद्याचे हाड) बळकट पण आखूड व रुंद झालेली असते. श्रोणी व मागील पाय पूर्णपणे ऱ्हास पावलेले किंवा अवशेषांगाच्या (लहान व अपूर्णपणे वाढलेल्या) रूपात राहिलेले असतात. पुच्छपक्ष क्षितिजसमांतर असतो.
स्पष्टपणे माशांसारखे दिसत असले आणि जलचर जीवनप्रणालीशी जुळवून घेण्यासाठी सांगाड्यात काही बदल झालेले असले, तरी सीटॅसियांच्या सांगाड्याची रचना मूलतः सस्तन प्राण्यांच्या सांगाड्यासारखीच आहे व त्यांचे पूर्वज मासे किंवा जलचर सरीसृप नसून बहुधा भूचर मांसाहारी अपरास्तनी असावेत या अपरास्तनींच्या दातांचे सर्व सामान्य सूत्र ३.१.४.३/३.१.४.३ आहे. सीटॅसियांचे आकारमान प्रचंड असून ते तलप्लावी किंवा समुद्रकिनाऱ्याजवळ राहणारे आहेत. डॉल्फिन सर्व समुद्रांत व काही मोठ्या नद्यांच्या मुखांत राहतात. सीटॅसिया इओसीनमध्ये अवतरले आणि मायोसीन, प्लायोसीन व प्लाइस्टोसीन काळांत ते विपुल व विविध प्रकारचे झाले होते.
(५) एडेंटाटा गण : या प्राण्यांची कवटी लांबट वा आखूड व खोलगट असते. मानेत सामान्यतः ७, क्वचित ९ वक्षात १२ ते २४ आणि कटीत ३ ते ९ मणके असतात. ग्लिप्टोडोंशिया गटात वक्षातील व इतर कित्येक मणके त्यांच्यावरील कंटकांसह जोडले जाऊन नलिका बनतात. हे प्राणी एकवारदंती आहेत परंतु टॅट्युसिया व ओरिक्टेरोपस वंशांतील दातांवरून एडेंटाटांचे पूर्वज द्विवारदंती असल्याचे दिसते. रीसेंट कालीन एडेंटाटांना सामान्यतः फक्त ४ ते १९ किंवा अधिकही दाढाच असतात. दाढांना दंतवल्क व दंतमूले नसतात आणि त्यांचे माथे जसजसे झिजतात तसतशा त्या आयुष्याभर वाढत असतात. दाढांवर पुष्कळदा उभे कटक असतात व ग्लिप्टोडोंशियांमध्ये ते ठळक असल्याने दाढा खांबाप्रमाणे दिसतात. जीवाश्मरूपातील प्राण्यांत कृंतक दात व सुळे होते आणि दंतिनावर दंतवल्क होते. एडेंटाटांचे मागील पाय सामान्यपणे अधिक बळकट असतात व कधीकधी ते पुढील पायांपेक्षा पुष्कळ लांब असतात पायांना ३ ते ५ बोटे असून टोकांच्या बोटांना चापट लांब नख्या असतात किंवा ती कधीकधी खुरासारखी असतात.
जनेनंद्रियांतील फरकांनुसार एडेंटाटा गणाचे (क) नोमॅर्थ्रा व (ख) झेनॅर्थ्रा हे उपगण करतात. नोमॅर्थ्रामध्ये मान व वक्ष यांमधील सांधा साधा असून त्यांचे थोडेच जीवाश्म आढळतात. ते पूर्व गोलार्धात सापडतात. झेनॅर्थ्रामध्ये मानेच्या पश्च व वक्षाच्या अग्र मणक्यांमध्ये अतिरिक्त सांधा असतो हिकॅनोडोंटा गटातील सर्व प्राण्यांना शृंगी वा अस्थिमय पट्टांचा बनलेला बाह्य कंकाल असतो परंतु ॲनिकॅनोडोंटा गटात बाह्य कंकाल निर्माण झालेला असतो किंवा नसतो.
पूर्व गोलार्धातील एडेंटाटांचे जीवाश्म इओसीन काळापासूनचे असून ते यूरोपात आढळतात, तर पश्चिम गोलार्धात दक्षिण अमेरिकेतील इओसीन कालीन खडकांमध्ये झेनॅर्थ्राचे विपुल जीवाश्म सापडले आहेत. ग्रॅव्हिग्रेडा (मोठे स्लॉथ) कुलातील प्राणी आताच्या अस्वलांच्या मानाने पुष्कळ अगडबंब व बेडौल होते. तसेच त्यांची शेपटी लांब होती. पॅटागोनियातील तृतीय व दक्षिण अमेरिकेतील प्लाइस्टोसीन कालीन खडकांमध्ये यांचे विपुल जीवाश्म आढळले आहेत. उत्तर अमेरिकेत यांचे प्लायोसीन व प्लाइस्टोसीन कालीन जीवाश्म सापडलेले आहेत. ⇨ आर्मंडिलो हे हिकॅनोडोंटा (झेनॅर्थ्रा उपगण) गटातील डॅसिपोडिडी कुलातील प्राणी आहेत. सध्या ते दक्षिण अमेरिकेच्या उष्ण व समशीतोष्ण कटिबंधीय भागांत राहत असून टॅटयुशिया या एकमेव वंशाचा प्रसार मध्य अमेरिकेपर्यंत (मेक्सिको व टेक्ससपर्यंत) झालेला आढळतो. यांचे जीवाश्म फक्त अमेरिकेत आढळले आहेत.
(६) रोडेशिया गण : [⟶ कृंतक गण]. यातील प्राणी सामान्यपणे लहान, शाकाहारी व बहुतेक बिळांत राहणारे, कधीकधी पोहणारे व आधारावर चढणारे आहेत. त्यांची कवटी बसकी असून मुस्कट बहुधा लांब व त्याचे पश्च टोक एकदम छाटले गेल्यासारखे दिसते. पुढच्या पायांना व मागच्या पायांना ५ किंवा ३ बोटे असून बोटांना नख्या असतात. दंतविन्यास अतिशय विशेषित असून दातांचे सूत्र, विशेषतः सिंप्लिसिडेंटाटांमध्ये २.०.३.३/१.०.३.२ ते १.२.२/१.०.२ असे बदलते. सुळे नाहीसे झाल्याने कृंतक व कपोल दातांमधील दंतावकाश मोठा असतो. कृंतकांचे माथे जसजसे झिजत जातात तसतसे ते आयुष्यभर वाढत जातात. उपदाढा अजिबात नसतातही व दाढा लहान, ब्रॅकिओडोंट आणि ब्युनोडोंट वा लोफोडोंट प्रकारचा माथा असलेल्या असू शकतात, तर काहींत त्या हिस्पोडोंट प्रकारच्या व प्राचिनाकार आणि खालून उघड्या व दंतमूलहीन असू शकतात. मूलतः वरच्या कपोल दातांना ३ तर खालच्यांना ४ दंताग्रे असून ती जोडली जाऊन कटक तयार होतात. कृंतक दातांची प्रतिष्ठापना केवळ डुप्लिसिडेंटाटा उपगणातच होते. उपदाढांमध्येच बदल घडून येतात. क्युव्हायनी उपकुलातील अलीकडच्या प्राण्यांमध्ये गर्भामध्ये कृंतक दात प्रतिष्ठापित होत असल्याचा पुरावा मिळाला आहे.
इतर सर्व सस्तन प्राण्यांच्या मानाने हल्लीच्या रोडेंशियांच्या वंशांची आणि जातींची संख्या जास्त आहे. त्यांच्या ९०० पेक्षा जास्त जाती माहीत असून त्यांपैकी पुष्कळ दक्षिण अमेरिकेत आहेत. यूरोप, आशिया व उत्तर येथील पुष्कळ वंश समान आहेत. खास आफ्रिकेतील म्हणता येतील असे पुष्कळ वंश आफ्रिकेत आहेत व त्याशिवाय आशिया व यूरोप येथे आढळणारे काही वंशही तेथे आहेत.
रोडेंशिया लहान व नाजूक चणीचे असले, तरी त्यांचे पुष्कळ जीवाश्म आढळतात. उत्तर अमेरिकेत हे पूर्व इओसीन काळात अवतरले. उत्तर इओसीन व मायोसीन काळांत हे युरोपात व उत्तर अमेरिकेत विपुल प्रमाणात होते. दक्षिण अमेरिकेत हे पूर्व तृतीय व प्लाइस्टोसीन काळांत अतिशय विपुल होते. यूरोप व दोन्ही अमेरिकेत येथील गुहांमध्ये साचलेल्या गाळात यांचे पुष्कळ जीवाश्म आढळले आहेत.
रोडेंशिया एम्. वेबर यांनी पुढील दोन उपगण केले आहेत : (क) सिप्लिसिडेंटाटा : यात कित्येक कुले व उपकुले आहेत आणि (ख) डुप्लिसिडेंटाटा : उपलब्ध असलेल्या माहितीप्रमाणे सर्व वर्गीकरणातील कदाचित हेच सर्वांत चांगले आहे, कारण इतरांनी सुचविलेल्या तीन वा चार गटांपैकी काहींमधील परस्परसंबंध ठरविणे अवघड आहे.
सिप्लिसिडेंटाटा : डुप्लिसिडेंटाटापेक्षा पुष्कळ मोठा गट असून त्यातील दंतविन्यास अधिक वैविध्यपूर्ण आहे. याउलट विविध प्रकारचे ससे ज्यात येतात त्या डुप्लिसिडेंटाटामध्ये वरच्या दोन कृंतक दातांना पूर्णपणे दंतवल्क असते आणि हे त्यांचे वैशिष्ट्य आहे. शिवाय कपोल दात हिप्सोडोंट प्रकारचे, दोन वा तीन प्रचिनांचे बनलेले, दंतमूलविहीन व वरच्या जबड्यात लांबीपेक्षा जास्त रुंद असे असतात.
सामान्यपणे या प्राण्यांचा जलदपणे विस्तृत प्रसार झाला त्यातल्या त्यात मायरिनी (घुशी व उंदीर) या उपकुलातील आणि हिस्ट्रिकॉइडिया (साळ) या कुलातील प्राण्यांची असा जलदपणे प्रसार होण्याची क्षमता सर्वकाळ विलक्षण असलेली दिसते. हे प्राणी पूर्व गोलार्धात मध्य तृतीय काळात अवतरले व जलदपणे पश्चिम गोलार्धात पसरले.
(७)अंग्युलेटा गण : [खुरी प्राणी⟶ अंग्युलेटा]. हे शाकाहारी, क्वचित सर्वभक्षी भूचर सस्तन प्राणी असून ते खुरांवर (बोटांवर) चालतात. ही चालण्याची तऱ्हा पादतलचारी, अर्धपादतलचारी ते अंगुलिचारी अशी विकसित झाली व त्यांच्या पावलाच्या पश्चपाद भागाचा (खूर व घोटा यांतील भागाचा) जमिनीला स्पर्श होईनासा झाला. मुळात कवटीचा माथा सपाट व बसका होता व अरुंद कवटीवर मध्यवर्ती कटक असे. कवटीचे विविध दिशांनी विशेषीकरण झालेले आहे. उदा., ललाटीय अस्थीत हवेच्या पोकळ्या निर्माण होणे किंवा आर्टिओडॅक्टिलांसारखी (समखुरी प्राण्यांप्रमाणे) शिंगे येणे. विस्तारलेली नासास्थी व नेत्रकोटरे आणि मुस्काटावरील पुढे आलेल्या भागाचे अस्थीभवन यांमुळे त्यांची चेहरेपट्टी वैशिष्ट्यपूर्ण अशी दिसते.
आदिम प्राण्यांत दंतविन्यास पूर्ण असून त्यांना जवळजवळ असलेले शंकूच्या आकाराचे कृंतक, सुळे व त्रिगुलिकायुक्त कपोल दात असतात. कृंतक दातांत परिवर्तन होऊन ते छिन्नीसारखे वा फावड्यासारखे, कधीकधी सुळ्यासारखे झालेले असतात अथवा काहींत ते आखूड झालेले असतात किंवा (विशेषतः वरचे) पूर्णपणे नाहीसेही झालेले असतात. सुळे अणकुचीदार किंवा लहान होतात. वरचे पूर्णपणे नाहीसे होतात, तर खालचे कृंतकासाऱखे होऊन कृंतक दातांचे कार्य करतात (उदा., रवंथ करणारे प्राणी). दोन वा जास्त दंताग्रे येऊन व कटक अथवा कोर निर्माण होऊन त्रिगुलिकायुक्त कपोल दात ब्युनोडोंट, लोफोडोंट व सोलेनोडोंट (कोरीच्या आकाराचे) प्रकारचे होतात. खालच्या दाढा वरच्यांपेक्षा लहान असतात. उपदाढांचा विकास बहुधा दाढांपेक्षा किंचित कमी झालेला असतो. कृंतक व सुळे यांना एकच दंतमूल असते, वरच्या दाढांना मुळात तीन दंतमुले असतात आणि खालच्या दाढांना दोन दंतमुले असून विभाजन होऊन त्यांची चार झालेली असतात. शिवाय मूळच्या ब्रॅकिओडोंट टप्प्याला त्यांच्यात हिप्सोडोंट व प्रचिनाकार होण्याची प्रवृत्ती आढळते.
हल्ली खुरी प्राणी जगभर विखुरलेले आहेत मात्र ऑस्ट्रेलियात वसाहत करणाऱ्यांनी ते आपल्याबरोबर नेले आहेत. त्यांचा तृतीय काळातील प्रसार जवळजवळ असाच होता.
खुरी प्राण्यांचे पुढील पाच उपगण पडतात : काँडिलार्थ्रा, लिटोप्टेर्ना, पेरिसोडॅक्टिला (विषमखुरी प्राणी), आर्टिओडॅक्टिला व अँब्लिपोडा. त्यांपैकी काँडिलार्थ्रा व लिटोप्टेर्ना हे आदिम प्रकारचे असून त्यांच्यात विविधता नव्हती किंवा त्यांचा विस्तृत प्रसार झालेला नव्हता. अँब्लिपोडांतील आदिम विशेषीकृत वैशिष्ट्यांच्या मिश्रणामुळे त्यांचे वर्गीकरणातील स्थान काहीसे अनिश्चित आहे. पेरिसोडॅक्टिला व आर्टिओडॅक्टिला यांचा (त्यातही विशेषतः आर्टिओडॅक्टिलांचा) त्यांच्या दीर्घ ऐतिहासिक काळात विस्तृत प्रसार झालेला आहे व त्यांच्यात पुष्कळ विविधता आलेली आढळते.
काँडिलार्थ्रा व अँब्लिपोडा हे इओसीनमध्ये तर लिटोप्टेर्ना पॅलिओसीन काळात निर्वंश झाले.
(क) काँडिलार्थ्रा उपगण : हा निर्वंश झालेला गट असून यूरोपातील इओसीनच्या प्रारंभीचे विखुरलेले अगदी थोडे जीवाश्म वगळता याचे जीवाश्म केवळ उत्तर अमेरिकेच्या पाश्चिम भागातील इओसीन कालीन खडकांत आढळतात. यांच्यापासून पूर्व इओसीन काळात पेरिसोडॅक्टिला तसेच, उत्तर इओसीन काळात आर्टिओडॅक्टिला व कदाचित दक्षिण अमेरिकेतील लिटोप्टेर्ना उपगणही निर्माण झाला.
यांच्या कवटीच्या व सांगाड्याच्या पुष्कळ रचना क्रिओडोंशियांप्रमाणे, तसेच आदिम प्रकारच्या पेरिसोडॅक्टिला व आर्टिओडॅक्टिला यांच्याप्रमाणे होत्या उदा., कवटी बाजूने बुटकी, सपाट व तिचा मध्यवर्ती कटक तसाच, मस्तिष्क-गुहा अगदी लहान, पूर्ण व जवळजवळ असलेल्या दातांचा संच, कृंतक व सुळे अगदी सारखे, कपोल दात ब्रॅकिओडोंट त्रिगुलिकायुक्त. त्यांच्या चालण्याचा टप्पा पादतलचारी व अर्धपादतलचारी अवस्थेचा होता व टोकाजवळील खूर चापट व पुष्कळदा दोन भाग झालेले होते आणि अरास्थी व प्रबाहु-अंतरास्थी ही हाडे भिन्न होती.
कपोल दात त्रिगुलिकायुक्त होते ते क्रमविकासात चार वा पाच गुलिकायुक्त झाले व उपदाढांची दाढा होण्याकडे प्रवृत्ती होती. या उपगणातील फॅनेकोडोंटिडी कुल अंगुलिचारी झाले होते. या संरचनात्मक साम्यांवरून काँडिलार्थ्रा व क्रिओडोंशिया हे समान पूर्वज शाखेपासून आले असले पाहिजेत, असे दिसते.
(ख) लिटोप्टेर्ना उपगण : हे प्राणी फक्त दक्षिण अमेरिकेतच होते. उत्तर इओसीन काळात हे अमेरिकेतील ब्युनोडोंट काँडिलार्थ्रापासून निर्माण झाले व त्यांना पेरिसोडॅक्टिलांसारखे दात प्राप्त झाले. सुरुवातीला दंतविन्यास पूर्ण होता, कृंतक दात लहान होण्याची प्रवृत्ती होती नमुनेदार ब्रॅकिओडोंट, क्वचित हिप्सोडोंट प्रकारचे कपोल दात विकासित झाले व खालच्या जबड्यातील अशा दातांना चार दंतमुले आणि काहीसे कोरीच्या आकाराचे कटक होते. ते अंगुलिचारी या चालण्याच्या अवस्थेत पोहोचले होते. बाजूचे खूर कमी होऊन तीन (किंवा क्वचित एकच) खूर त्यांना होते व त्यांपैकी कडेचे खूर रुंद असत. त्यांच्या पायांची हाडे पुष्कळशी आदिम पेरिसोडॉक्टिलांच्या पायांच्या हाडांसारखी होती. हा उपगण प्लाइस्टोसीन काळात निर्वंश झाला.
(ग) अँब्लिपोडा उपगण : हा उपगण केवळ इओसीन काळात आढळतो. त्यांचे वर्गीकरणातील स्थान निश्चितपणे माहीत नाही. या उपगणातील प्राण्यांची मस्तिष्क-गुहा सर्व खुरी प्राण्यांमध्ये सर्वांत लहान आहे. पँटोलँब्डिडी या आदिम कुलाचा प्रारंभ लहान प्राण्यांनी झाला त्यांची कवटी मांसाहारीपर्माणे बसकट, तर पायांना पाच खूर होते आणि ते पादतलचारी ते अर्धपादतलचारी होते. त्यांचा दंतविन्यास पूर्ण होता. कृंतक व सुळे यांची सर्वसाधारण अशी वाढ झालेली असे व कपोल दातांवर V आकाराचे कटक असत.
कॉरिफोडोंटिडी कुलातील प्राणी मोठे होते. त्याच्या कपोल दातांवरील दंताग्रे V आकाराच्या कटकाने जोडलेली होती व सुळे हत्तीच्या दातांप्रमाणे अतिशय लांब होते. कवटी इतर शरीराच्या मानाने खूप मोठी होती. ते अंगुलिचारी होते व पायांच्या रचनेवरून ते हळू चालणारे होते, असे दिसते.
डायनोसेरॅटिडी कुलात अधिक विविधता होती. खालचे कृंतक दात लहान झालेले होते आणि वरचे सुळे हत्तीच्या दातांसारखे व खालचे सुळे लहान होते. कवटीच्या ललाटीय भागावर शिंगांसारख्या रचना असत. या कुलातील काही प्राणी प्रचंड आकारमानाचे होते उदा., लोक्झोलोफोडॉन (वायोमिंग येथील इओसीन कालीन जीवाश्म) वंशातील प्राण्यांची उंची २ मी. व लांबी ४ मी. होती. त्यांचे पाय बळकट आणि पादास्थी (पायांची हाडे) प्रोबॉसिडियाप्रमाणे (हत्तीप्रमाणे) होत्या. अशा प्रकारे पेरिसोडॅक्टिला, ऑर्टिओडॅक्टिला व प्रोबॉसिडिया यांच्यातील कवटीची सांगाड्याची आदिम वैशिष्ट्ये तसेच विशेषीकृत वैशिष्ट्ये या उपगणात एकवटलेली आहेत. हा उपगण केवळ इओसीनमधील अल्पकाळातच आढळत असला, तरी याच्या उत्तर अमेरिका, यूरोप व मंगोलिया येथील जीवाश्मांवरून त्या अल्पकाळातच त्यांचा पुष्कळ प्रसार झालेला होता, असे दिसून येते.
(घ) पेरिसोडॅक्टिला उपगण : हल्ली या उपगणाचे टॅपिर, ऱ्हिनोसेरॉस व ईक्वस (घोडा) हे तीनच वंश (व त्यांना जवळचे असलेले परिवर्तित प्राणी) आहेत. त्यांच्या कवटीवर कानशीलाच्या भागात कटक असतो. अनुकपाल (कवटीचा पश्चकपालास्थीय भाग) मागील बाजूस विषम उताराचे असते आणि अस्थिकंद आडव्या दिशेत फुगीर असतो. ईंक्वस वंश वगळता इतरात नेत्रकोटरे पश्च दिशेने उघडी असतात ईक्वस वंशात नेत्रकोटरांभोवती हाड असते. ऱ्हिनोसेरॅटिडी व टिटॅनोथेरिआयडी यांच्यात नाकावर शिंगे वा बाहेर आलेला अस्थिमय भाग असतो.
यांच्या कायमच्या दातांचे नमुनेदार सूत्र ३.१.४.३/३.१.४.३ असे असून ते इओसीन कालीन सर्व प्राण्यांत आढळते. कृंतक व वरचे सुळे (क्वचित खालचेही) तसेच सर्वांत पुढची उपदाढ हे दात लहान होतात किंवा त्यांचे पूर्णतया दमन होते. आदिम ब्रॅकिओडोंट प्रकारच्या दातांचे विशेषीकरण होऊन ते हिप्सोडोंट प्रकारचे व प्रचिनाकार होतात आणि चतुर्गुलिकायुक्त माथा क्वचित ब्युनोडोंट प्रकारचा राहतो परंतु सामान्यतः दंताग्रे एकत्र येऊन तो लोफोडोंट प्रकारचा होतो.
कशेरुक-अक्षाचे मानेतील ७, पृष्ठकटीय २० ते २६, त्रिकास्थीय ५ ते ६ आणि शेपटीतील १३ किंवा जास्त मणके असतात. भुजास्थी आखूड व बळकट असून प्रबाहू व प्रबाहू- अंतरास्थी वेगळ्या असतात किंवा त्या जोडल्या जाण्याची प्रवृत्ती असते. दूरस्थ मणिबंधास्थी (मनगटाचे हाड) बाजूला सरकून व ती जास्त व्यवस्थित झाल्याने अतिशय बळकट अशी मनगटांची रचना निर्माण होते. उर्वस्थीला विकसीत झालेले तिसरे उरुकूट (मांडीच्या हाडाच्या वरच्या टोकाशी असलेला प्रवर्ध म्हणजे वाढ) असते व आर्टिओडॅक्टिलातील ऊर्वस्थीच्या तुलनेने यांचे हे वैशिष्टय आहे. ज्यांचा मधला खूर प्रमुख नसतो त्या प्राण्यांतील अंतर्जघास्थी (गुडघा व घोटा यांच्यामधील आतले व मोठे हाड) व बाह्य जंघास्थी (नडगीचे बाहेरचे व लहान हाड) वेगळ्या असतात परंतु ईक्वीडीमधील प्रगत प्राण्यांत जेव्हा मधला खूर सर्वांत प्रमुख होतो तेव्हा बाह्य जंघास्थी अंतर्जघास्थीच्या बरोबरच वाढतात व बाह्य जंघास्थी लहान कपचीसारखी होते. पुढील पायांपेक्षा मागील पायांतील बाजूचे खूर पूर्णतया लहान झालेले असतात आणि ईक्विडीच्या अगदी प्रगत प्राण्यांत एकखुरी टप्पा विशेष दिसून येतो. अशा प्रकारे शरीराचे एकूण वजन मधल्या खुरावर पडते.
खुरांची संख्या घटून जरी उत्तर इओसीन काळातच पुढील पायांना चार व मागील पायांना तीन खूर होते, तरी हल्लीच्या टॅपिर या अपरिवर्तनशील वंशात ती तशीच टिकून राहिली आहेत [⟶ टॅपिर]. टॅपिरिडी कुलाचे सर्वात आधीचे जीवाश्म यूरोपातील आणि उत्तर अमेरिकेतील इओसीन कालीन खडकांत सापडले आहेत. या अपरिवर्तनशील कुलाचा प्लायोसीन काळापर्यंतचा उत्तर गोलार्धातील इतिहास गतिमान नसला, तरी दीर्घकालीन आहे. प्लाइस्टोसीन काळापासून या कुलाचे प्राणी अमेरिका व आग्नेय आशियातील केवळ उष्ण कटिबंधीय वनांमध्ये असून आता यांचा टॅपिर हा एकमेव वंश राहिलेला आहे. यांच्या पायांचे जसे परिवर्तन झाले नाही, तसेच यांच्या दातांतही परिवर्तन झालेले नसून बुटक्या माथ्याच्या कपोल दातांवर साधेच कटक आहेत.
ऱ्हिनोसेरॅटिडी कुलाचे सर्वांत आधीचे जीवाश्म यूरोप व उत्तर अमेरिकेतील मध्य इओसीन कालीन खडकांत सापडले असून मायोसीन, प्लायोसीन व प्लाइस्टोसीन काळांत त्यांचा उत्तर गोलार्धात विस्तृत प्रसार झाला होता. हल्ली हे प्राणी केवळ मध्य आफ्रिका. भारत सुंदा बेटे (आग्नेय आशिया) येथील कमी उंचीच्या दलदली भागांत आढळतात आणि तेथे त्यांचा ऱ्हिनोसेरॉस हा एकमेव वंश आढळतो [⟶ गेंडा]. या कुलातील प्राण्यांचे शरीर बोजड असून त्यांचे पाय आखूड व मजबूत असतात. त्यातल्या त्यात प्रगत प्राण्यांच्या पुढील पायांना तीनच खूर असतात. कवटी मोठी असून कपाळ व नाक या भागांत बहुधा एकामागे एक अशी परंतु डायसेरॅथेरियम वंशाप्रमाणे क्वचित एकमेकांच्या बाजूला अशी दोन शिंगे असतात. सुरुवातीच्या प्राण्यांत दंतविन्यास पूर्ण असतो. कृंतक व सुळे नाहीसे होण्याची प्रवृत्ती असून दाढा व उपदाढा जवळजवळ सारख्या असतात आणि त्यांच्यावर मऊ वनस्पती-अन्नाला अनुरुप अशा साध्या दुमडी असतात.
दात व पाय यांत झालेले बदल सर्वांत ठळकपणे ईक्विडी कुलात दिसून येतात आणि पेरिसोडॅक्टिलांपैकी या कुलाच्या दातांची व पायांची माहिती सर्वात पूर्ण तपशीलवारपणे माहिती झालेली आहे [⟶ घोडा]. इनोसीनमध्ये अवतरल्यापासून या कुलातील प्राण्यांत झालेल्या क्रमविकासाचे विविध टप्पे कोष्टकात (पृष्ठ ९३०) वंशांच्या क्रमाने दिलेली आहेत. त्यांचे जीवाश्म ज्या ज्या ठिकाणी सापडले आहेत तेथील निक्षेपांचे सहसंबंध ठरविण्यासाठी क्रमविकासाचे विविध टप्पे स्पष्ट करणारे हे वंश अतिशय उपयुक्त ठरले आहेत. ईक्वस वंश भारतामध्ये मध्य (किंवा कदाचित पूर्व) प्लायोसीनमध्ये निर्माण झाला, उत्तर प्लायोसीनमध्ये यूरोपात आणि प्लायोसीनच्या शेवटी किंवा प्लाइस्टोसीनच्या सुरुवातीस उत्तर अमेरिका व आफ्रिका या भागांत त्याचा प्रसार झाला. अशा प्रकारे प्लाइस्टोसीन काळात ऑस्ट्रेलिया वगळता सर्व जगभर याचा प्रसार झाला होता. त्यामुळे हा वंश अमेरिका व आफ्रिकेच्या प्रदेशांत प्लायोसीन-प्लाइस्टोसीन कालीन संक्रमणाचा निर्देशक मानला जातो.
ईक्वाइन सस्तन प्राण्यांच्या भूवैज्ञानिकप्रसार व जातिवृत्त
| उत्तर आफ्रिका | यूरोप | आशिया | उत्तर अमेरिका | दक्षिण अमेरिका | |
| रीसेंट | ईक्कस | ईक्कस | ईक्कस | ईक्कस (यूरोपातून आला) | ईक्कस (यूरोपातून आला) |
| प्लाइस्टोसीन (प्रलय जडोढ) | ईक्कस | ईक्कस | ईक्कस | ईक्कस | हिप्पिडियम (ईक्कस) |
| उत्तर प्लायोसीन | ईक्कस | ||||
| मध्य प्लायोसीन | हिप्पेरिऑन | हिप्पेरिऑन | ईक्कस | ||
| पूर्व प्लायोसीन | हिप्पेरिऑन | हिप्पेरिऑन अँकिथेरियम | प्रोटोहिप्पस प्लायोहिप्पस मेरिकहिप्पस | ||
| उत्तर मायोसीन | अँकिथेरियम | निओहिप्पेरिऑन मेरिकहिप्पस | |||
| मध्य मायोसीन | अँकिथेरियम | मेरिकहिप्पस पॅरॅहिप्पस हायपोहिप्पस | |||
| पूर्व मायोसीन | मेसोहिप्पस मायोहिप्पस | ||||
| ऑलिगोसान | मेसोहिप्पस मायोहिप्पस | ||||
| उत्तर इओसीन | इपिहिप्पस | ||||
| मध्य इओसीन | ओरोहिप्पस | ||||
| पूर्व इओसीन | इओहिप्पस |
ईक्वस वंशाचे स्तरवैज्ञानिक महत्त्व पुष्कळ असल्याने त्यादृष्टीने त्याच्या प्रारंभकालाविषयी मतभेद आहेत. भारतात व चीनमध्ये यांचे प्लोयोसीन कालीन, यूरोपात उत्तर प्लोयोसीन व उत्तर अमेरिकेत प्लायोसीन अखेरच्या वा प्लाइस्टोसीन प्रारंभीच्या काळातील जीवाश्म मिळाल्याचा दावा केला जातो. हे मतभेद मुख्यतः खरा ईक्वस वंश नेमका केव्हा अस्तित्वात आला, या मुद्याविषयी आहेत. पायांचा क्रमविकास विचारात घेतला, तर हेच पुढील शब्दात सांगता येते. दोन आणि चार क्रमांकांचे खूर जमिनीवर टेकण्याचे केव्हा थांबले, तसेच तीन क्रमांकाचा खूर हा पायाचे एकमेव टोक केव्हा झाला, हे ठरविणे अगदी सोपे असू शकेल असे नाही. शिवाय तरूण प्राण्यांच्या दाढा या पूर्णपणे म्हाताऱ्या प्राण्यांच्या दाढांसारख्या असण्याची शक्यता निदान काही अंशी कमी असणार. कारण त्याच्या आयुष्यभरच्या काळात दाढांवर दलनाचा परिणाम झाला असणार आणि त्यामुळे निश्चित निर्णय घेणे काहीसे अवघड होणार.एक लक्षात ठेवण्यासारखी गोष्ट म्हणजे ईक्विडी कुलाचा पूर्ण क्रमविकास उत्तर अमेरिकेत झाला व तेथूनच हे प्राणी यूरोप, आशिया व दक्षिण अमेरिका या प्रदेशांत गेले असले, तरी उत्तर अमेरिकेत ईक्वसवंश प्लइस्टोसीनच्या शेवटी लुप्त झाला होता आणि यूरोपातून आलेल्या वसाहतवाल्यांनी हा प्राणी पुन्हा उत्तर अमेरिकेत आणला.
कॅलिकोथेरिआयडी व टिटॅनोथेरिआयडी या कुलांतील प्राणी विलक्षण व थोराड चणीचे होते त्यांचे पाय आखूड व मजबूत होते. दातांचा विकास थोडाच झाला होता व ते मऊ वनस्पती खाण्याच्या दृष्टीने सोयीचे होते. टिटॅनोथेरिआयडी कुलातील प्राण्यांत दोन शिंगे होती. मुख्यत्वे उत्तर अमेरिकेत इओसीन व ऑलिगोसीन काळांत त्यांची भरभराट झाली. कॅलिकोथेरिआयडी कुल उत्तर अमेरिका व यूरोपमध्ये तृतीय काळात, भारत व चीनमध्ये प्लायोसीन आणि युगांडात प्लाइस्टोसीन काळात होते. प्लाइस्टोसीन काळात ते निर्वंश झाले.
(च) आर्टिओडॅक्टिला उपगण : आता उतरती कळा आलेल्या पेरिसोडॅक्टिलांच्या उलट आर्टिओडॅक्टिला हा विविधता, विपुलता व प्रसार या दृष्टीने हल्लीच्या ॲंग्युलेटांपैकी सर्वांत प्रमुख गट आहे. त्याच्या खुरांची संख्या सम असते, हे त्यांचे वैशिष्ट्य असून तीन व चार क्रमांकांचे खूर सारखे वाढलेले असतात व बाजूचे बारीक किंवा अल्पविकसित असतात. कवटी लांब असून ललाटास्थी नेहमीच कवटीचा एक प्रमुख भाग असते. ललाटास्थीवरील शिंगांमुळे वा शिंगांचे गाभे यांमुळे कवटीला वैशिष्ट्यपूर्ण रुप आले आहे.
दंतविन्यास सामान्यपणे पूर्ण असून दातांचेसर्वसाधारण दंतविन्यास सामान्यपणे पूर्ण पूर्ण असून दातांचे सर्वसाधारण सूत्र ३.१.४.३/३.१.४.३ असे असते. रंवथ करणाऱ्या प्राण्यांत दंतावकाश विशेष मोठा असतो व तो जबडा लांबट झाल्याने किंवा सुळे व उपदाढा यांचे दमन झाल्याने निर्माण झालेला असतो.
खालच्या जबडयापेक्षा वरच्या जबड्यातील कृंतकांचे व सुळ्यांचे लघुभवन (बारीक होणे), परिवर्तन व दमन ही अधिक लवकर होतात. सर्व सुरुवातीच्या व आदिम प्राण्यांचे दात ब्युनोडोंट प्रकारचे असतात परंतु रवंथ करणाऱ्या प्राण्यांमध्ये ते प्रचिनाकार असतात. पाच वा सहा गुलिकायुक्त दात प्रगत प्राण्यांमध्ये चार गुलिकायुक्त झालेले असतात. ब्युनोडोंट दात ब्युनो-सेलेनोडोंट होतात व सेलेनोडोंट दंताग्रे V अकाराची व कोरीच्या आकाराची होतात. खालच्या दाढा वरच्या दाढांपेक्षा अरूंद असतात.
भुजास्थी आखूड व बळकट असते तर प्रबाहू व प्रबाहु-अंतरास्थी प्रगत प्राण्यांत सांधल्या जाण्याची प्रवृत्ती आढळते. बाजूचे खूर ऱ्हास पावण्याची प्रवृत्ती सर्व आर्टिओडॅक्टिलांध्ये आढळते आणि प्रगत प्राण्यांमध्ये हे खूर दमन तीक्ष्ण झालेल्या टोकांसारखे लोंबत असतात. फक्त ॲग्रिओकीरिडी व अँग्रॅकोथेरायडी कुलांत पहिली करभास्थी (तळव्याचे हाड) अवशेषांग रूपात टिकून राहिलेली असते. सुइडी व हिप्पोपोटॅमिडी कुलांमध्ये चार करभास्थी टिकून राहिलेल्या आहेत. पेरिसोडॅक्टिलांच्या उलट आर्टिओडॅक्टिलांमध्ये ऊर्वस्थीवर तिसरे उरुकूट नसते. प्रगत प्राण्यांत मागील पायांच्या बाह्य जंघास्थी अंतर्जंघास्थीशी जोडली जाण्याची प्रवृत्ती असते आणि पुढील पायांच्या खुरांप्रमाणे खूर बारीक होतात.
दंतविन्यासानुसार आर्टिओडॅक्टिलांचे पुढील तीन गट मानतात. ब्युनोडेंशिया : यांत डुकरे व हिप्पोपोटॅमस येतात ब्युनो-सेलेनो-डोंशिया : इतर दोन गटांतील काही प्राण्यांशी नाते असलेली काही कुले यात येतात व सेलेनोडोंशिया : यात १७ ते १८ कुले असून त्यांपैकी रवंथ करणारे प्राणी सर्वांत प्रमुख आहेत.
ब्युनोडोंशिया : यांच्यापैकी सुइडी कुल यूरोपात इओसीनमध्ये अवतरले व पूर्ण इओसीनमध्ये मायोसीनमध्ये पूर्व गोलार्धात त्यातील प्राणी विपुल होते व त्यांचा विस्तृत प्रसार झाला होता परंतु त्यामानाने अमेरिकेत ते तेवढे विपुल नव्हते. हल्ली ते युरोप, आशिया, आफ्रिका व अमेरिका या प्रदेशांतील मूळचे प्राणी आहेत. हिप्पोपोटॅमिडी कुलातील प्राणी काहीसे अगडबंब व बेडौल चणीचे तसेच अपरिवर्तनशील आहेत. आशिया, आफ्रिका व यूरोप येथे यांचे प्लायोसीन व प्लाइस्टोसीन कालीन जीवाश्म आढळले असून हल्ली आफ्रिकेच्या उष्ण कटिबंधीय भागात याचा हिप्पोपोटॅमस हा एकमेव वंश राहिला आहे [⟶ पाणघोडा]. लेप्टोकीरिडी या छोट्या कुलाचे जीवाश्म डकोटातील ऑलिगोसीन कालीन खडकांशिवाय इतरत्र थोडेच आढळले आहेत.
ब्युनो-सेलेनोडोंशिया : अँथ्रॅकोथेरायडी, ॲनोप्लोथेरायडी व डायकोब्युनिडी ही यातील तीन कुले आहेत. हा आदिम आर्टिओडॅक्टिलांचा गट असून यांचा इतर दोन गटांशी या ना त्या प्रकारे नातेसंबंध आहे. अशा प्रकारे अँथ्रॅकोथेरायडी कुलातील प्राण्यांचे सुइडी कुलातील सुरुवातीच्या प्राण्यांशी जवळचे संबंध असून दोन्ही प्राण्यांची पूर्वजपरंपरा बहुधा समान असावी. त्यांचे आकारमान डुकरापासून ते गेंड्याएवढे (ऱ्हेनोसेरॉसएवढे) होते. यूरोपात इओसीन व मायोसीन काळांत हे प्राणी विपुल होते परंतु त्यामानाने उत्तर अमेरिका, आफ्रिका व भारत येथे थोडेच जीवाश्म आढळतात.त्यां शंक्वाकार कृंतक दात चमसाकृती (चमच्याच्या आकाराचे) झाले व कपोल दातांना V आकाराची दंताग्रे येत गेली. सुळे मजबूत होते. पायांना चार खूर होते.
ॲनोप्लोथेरायडी कुलातील प्राण्यांच्या कवटीस आखूड मुस्कट असले, तरी ती कॅमेलसमधील (सेलेनोडोंशियातील) कवटीसारखी होती. कृंतक दात व सुळे तीक्ष्ण होते व कपोल दात सेलेनोडोंटासारखे दिसत. शरीर मजबूत असून आखूड व मजबूत पायांना टोकांशी नख्या असलेली चार बोटे असत. ते सर्वसाधारणपणे अंग्युलेटांपेक्षा अधिक मांसाहारींसारखे दिसत असत. ते दलदलीच्या भागात राहत व बहुधा त्यांच्या बोटांमध्ये पटलासारखी जाळीदार रचना असावी. हा गट मुख्यत्वे केवळ यूरोपामध्ये इओसीन व ऑलिगोसीन काळांत होता.
डायकोब्युनिडी कुलातील प्राण्यांच्या पायांना चार खूर होते. मात्र त्यांची कवटी पुष्कळशी ट्रॅग्युलस वंशाच्या रवंथ करणाऱ्या प्राण्यांच्या कवटीसारखी होती. उपदाढा साध्या होत्या व काही दाढांची अग्रे V आकाराची होती. आर्टिओडॅक्टिलांतील इतर सर्व कुलांच्या उलट या कुलातील प्राण्यांच्या दातांना सहा दंताग्रे होती. मात्र एकूण हा गट सुइडीसारखा आहे. या कुलाचे जीवाश्म केवळ उत्तर अमेरिका व युरोप येथील इओसीन व ऑलिगोसीन कालीन खडकांत आढळले आहेत.
सेलेनोडोंशिया : यातील आधीच्या तीन कुलांपैकी ॲग्रिओकीरिडी (म्हणजे ऑरिओडोंटिडी) कुल उत्तर अमेरिकेत इओसीन ते प्लायोसीन काळात होते याच्यातील प्राण्यांच्या पायांना चार खूर असत आणि पुढील पायांवर अवशेषी रूपातील आंगठा हे आदिम वैशिष्टय टिकून राहिले होते.
सीनोथेरायडी : हे आधीचे व आदिम कुल असून यातील प्राण्यांच्या पायांना चार खूर असत. हे प्राणी आकारमानाने सशाएवढे होते व ते कळप करून राहत असत. त्यांचे जीवाश्म उत्तर इओसीन ते मायोसीन काळातील असून ते युरोपात आढळतात.
झिफोडोंटिडी कुलातील प्राण्यांचे पाय लांब व बारीक होते व त्यांना दोन खूर असत. हे प्राणी ट्रॅक्विलिडीतील रवंथ करणाऱ्या प्राण्यांसारखे व हरिणांसारखे होते. यांचे जीवाश्म केवळ यूरोपात उत्तर इओसीन कालीन खंडकांमध्ये सापडले आहेत. कॅमलिडी गटातील प्राण्यांची चेहेरेपट्टी वैशिष्ट्यपूर्ण होती व त्यांची कवटी बाजूने पुष्कळशी घोड्यांच्या कवटीसारखी दिसे. त्यांना शिंगे नसत व ते रवंथ करणारे होते. त्यांचे पाय लांब व बारीक असून त्यांना दोन ते चार खूर असून प्रगत प्राण्यांत बाजूच्या खुरांचा पूर्णपणे क्षय झालेला होता त्याच्या दातांचे सूत्र ३.१.१.४-२.३/३.१.४-१.३ असे असून वरच्या कृंतकांची व खालच्या उपदाढांची लहान होण्याची प्रवृत्ती दिसते. सुळा व पहिली उपदाढ यांच्यात कधीकधी पहिल्या उपदाढेनंतर दंतावकाश असे. मणिबंधास्थी व गुल्फास्थी (घोट्याचे हाड) या स्वतंत्र असतात हे त्याचे वैशिष्ट्य होय कॅमलिडीतील प्राणी उत्तर अमेरिकेत इओसीनमध्ये अवतरले व मायोसीनपर्यंत तेथेच राहिले. प्लायोसीन काळात ते उत्तर आफ्रिका व आशियात गेले. यांच्या जीवाश्मांच्या नोंदी जवळजवळ परिपूर्ण आहेत. आता असलेल्या दोन वंशांपैकी कॅमेलस (उंट) आशिया व उत्तर आफ्रिकेत असून ऑकॅनिया वंश दक्षिण अमेरिकेत आहे.
ट्रॅग्युलिडी (माउस डियर) कुलातील प्राणी लहान, शिंगे नसलेले आदिम प्रकारचे रवंथ करणारे प्राणी होते. डायकोब्युनिडी कुल त्यांचे पूर्वज असून त्यांच्या गेटोसिनी उपकुलापासून कॅव्हिकॉर्नियातील सर्व्हिडी गट निर्माण झाला. यूरोप, आशिया व आफ्रिका येथे यांचे ऑलिगोसीन ते पूर्व प्लायोसीन कालीन जीवाश्म आढळले आहेत. प्लायोसीनपासून हे प्राणी दक्षिण आशिया व पश्चिम आफ्रिका या भागांत राहत आहेत.
हायपरट्रॅग्युलिडी या केवळ उत्तर अमेरिकेत आढळणाऱ्या छोट्या गटाला आता स्वतंत्र कुलाचा दर्जा देण्यात आला आहे. कारण ट्रॅग्युलिडीतील प्राण्यांच्या दातांशी तुलना केल्यास यांच्या कपोल दातांमध्ये काही रचनात्मक फरक आहेत.
सर्व्हिकॉर्निया व कॅर्व्हिकॉर्निया : विविध प्रकारची हरिणे, शेळ्या, मेंढ्या, गायी यांच्यात पुष्कळ फरक असून काही तज्ञ यांची वेगवेगळी कुले करतात परंतु येथे त्यांची काही समाईक वैशिष्ट्ये विचारात घेऊन त्यांचे पुढील दोन स्थूल गट केले आहेत सर्व्हिकॉर्निया व कॅव्हीकॉर्निया.
सर्व्हिकॉर्निया : हा लांबट कवटी असलेल्या हरिणांचा गट आहे. कवटी बरीच चापाकार असून सुरूवातीच्या प्राण्यांना शिंगे नव्हती परंतु मध्य मायोसीन काळापासून त्यांना शिंगे येऊ लागली आणि उत्तर प्लायोसीनपासून त्यांच्या शिंगांचे आकारमान आणि त्यांना फुटलेले फाटे हे त्यांचे ठळक वैशिष्ट्य झाले. सामान्यतः नरांना मृगशिंगे [⟶ मृगशृंगे आणि शिंगे] असतात परंतु रेनडियरांतील माद्यांनाही ती असतात. मृगशृंगांना वर्गीकरणाच्या दृष्टीने काही महत्त्व नाही. सर्व्हिकॉर्नियांच्या दंतविन्यासाचे सर्वसाधारण सूत्र ०.०-१.३.३/३.१.३(४).३ असे असून सामान्यपणे त्यांना वरचे कृंतक दात नसतात काही प्रगत प्राण्यांत सुळेही नष्ट झालेले आढळतात. कपोल दात ब्रॅकिओडोंट विशेषतः सेलेनोडोंट प्रकारचे असून त्यांच्यावर माथ्यावर जाड व चुरडलेले दंतवल्क असते. उपदाढा सापेक्षतः लांबट असतात. पायांतील २ ते ४ क्रमांकाच्या पश्चपादास्थी सांधल्या जाऊन बनलेली नालास्थी असते व जिराफिनी कुल वगळता इतरांत बाजूचे खूर आखूड असतात.
सर्व्हिकॉर्निया : ऑस्ट्रेलिया वगळता सर्व जगभर पसरले होते. यूरोपातील सर्व्हिकॉर्नियांचा प्रारंभ पूर्व मायोसीन काळात झालेला आहे. उत्तर मायोसीन व प्लायोसीन काळांमध्ये हरिणांचा पूर्व गोलार्ध व उत्तर अमेरिकेत विस्तृत प्रसार झाला होता आणि प्लाइस्टोसीन व रीसेंट काळात त्यांची जास्तीत जास्त भरभराट झाली होती.
प्रोटोसेरॅटिनी हा सर्व्हिकॉर्नियांचा सर्वांत आधीचा गट असून तो उत्तर इओसीनमध्ये (किंवा ऑलिगोसीनच्या प्रारंभी) अवतरला व तो मायोसीनपर्यंत होता त्याचे जीवाश्म फक्त उत्तर अमेरिकेत आढळले आहेत. त्यातील प्रगत प्राण्यांच्या लांबट चापट कवटीवर अस्थिमय प्रवर्ध होते. काहींना वरचे कृंतक दात नव्हते. त्यांना चार खूर होते व बाजूचे खूर बहुधा दुर्बल होते. बहुतकरून यांच्यापासून शिवाथेरायनी कुलाचे प्राणी अस्तित्वात आले असावेत.
मॉस्किनी हा बहुधा सर्व्हिकॉर्नियांतील सर्वांत छोटा गट असून त्यातील प्राण्यांना शिंगे नसतात व त्यांचे वरचे सुळे वाकड्या तलवारीसारखे असतात. त्यांचे जीवाश्म भारतातील प्लायोसीन कालीन शिवालिक खडकांत प्रथम आढळले आहेत. या गटाचे हल्ली मॉस्कस व हायड्रोपोटस हे दोनच वंश असून ते मध्य आशियातील डोंगराळ भागांत आढळतात.
सर्व्ह्युलिनी गटाच्या प्राण्यांना शिंगे नसतात किंवा फक्त नरांना लांब वृंतावरील (देठासारख्या भागावरील) आखूड मृगशृंगे असतात वरचे सुळे अतिशय बळकट असून ते खालच्या जबड्यावर आलेले असतात. मायोसीन काळापासूनचे यांच्या जीवाश्मांचे कित्येक वंश आढळले असून हल्ली या गटाचे प्राणी दक्षिण आशियात आहेत.
सर्व्हिनी उपकुलातील प्राण्यांची मृगशृंगे आखूड वृंतावर असून त्यांचे पुष्कळ विविध प्रकार आहेत. वरचे सुळे कमजोर असतात किंवा ते नसतातही. कपोल दात ब्रॅकिओडोंट प्रकारचे असतात. हल्ली हे प्राणी उत्तर गोलार्धात व दक्षिण अमेरिकेत राहात आहेत. त्यांचे प्लायोसीन काळापासूनचे जीवाश्म सापडले आहेत. प्लायोसीन व प्लाइस्टोसीन काळात त्यांचा विस्तृत प्रसार झाला होता. रेनडियर व रँगिफर यांच्या मृगशृंगांवरील फाटे जटिल असून हे प्राणी उत्तर गोलार्धातील थंड प्रदेशांमध्ये राहात आहेत. पुराणाश्मयुगीन (मॅग्डेलेनियन टप्पा) ऐतिहासिक वसाहतींमध्ये यांचे विपुल जीवाश्म सापडले आहेत. मेगॅसेरॉस हायबर्निकस या आयरिश एल्कच्या जातीचे प्राणी या उपकुलातील सर्वांत मोठे प्राणी असून त्यांच्या मृगशृंगाचा एका टोकापासून दुसऱ्या टोकापर्यंतचा व्याप ३ मी. पेक्षा जास्त होता.
बहुधा प्रोटोसेरॅटिनींपासून आलेल्या शिवाथेरायनी गटाचे सर्वांत चांगले माहीत असलेले पूर्व प्लायोसीन कालीन जीवाश्म भारतात आढळतात परंतु चीन व एड्रीॲनोपल (तुर्कस्तान) येथेही यांचे थोडे जीवाश्म सापडले आहेत. त्यांचे शरीर बळकट चणीचे असून त्यांच्या कवटीचा माथा हवायुक्त पोकळी असलेला असून त्यांच्या ललाटीय भागावर अल्पसे फाटे असलेली पट्टासारखी दोन मृगशृंगे होती व कधीकधी अग्र ललाटीय भागावर प्रवर्धांची दुसरी जोडी असे.
जिराफिनी गटातील प्राण्यांची कवटी लांबट असून तिची हाडे हवायुक्त पोकळ्या असलेली असतात. त्यांना शिंगे नसतात किंवा ललाटीय भागावर आखूड अस्थिमय स्कंधांची (फुगीर भागांची) जोडी असते अथवा नसते. कपोल दात बुटके, साधे, दबलेले पण कधीकधी जटिल व खोल असतात. उदा., कॅमेलोपारडॅलिस (जिराफ). या गटाचे जीवाश्म पूर्व प्लायोसीन काळापासून असून ते दक्षिण यूरोप, पर्शिया, भारत व चीन येथे सापडतात. हल्ली या गटाचे प्राणी मध्य व दक्षिण आफ्रिकेत आहेत.
अँटिलोकॅप्रिडी हा सर्व्हिकॉर्निया व कॅव्हीकॉर्निया यांच्याशी संबंधित असलेला गट असून त्यांना चुकून अँटिलोप (हरीण) म्हणतात. त्यांचे दात अँटिलोपांसारखे हिप्सोडोंट प्रकारचे असतात मात्र शिंगे भिन्न प्रकारची असून त्यांच्या शिंगांचा गाभा बोव्हिड आर्टिओडॅक्टिल (उदा., गाय, म्हैस) प्राण्यांप्रमाणे अस्थिमय असतो. यातील प्राणी सशाएवढे लहान होते. हा अलग असा लहान गट असून ऑलिगोसीन काळापासून केवळ उत्तर अमेरिकेत होता. बहुधा या प्राण्यांपासून कॅव्हिकॉर्निया प्राणी उदयास आले असावेत.
कॅव्हिकॉर्निया (पोकळ शिंगे असलेले रवंथ करणारे प्राणी). अंग्युलेटातील हा सर्वांत विपुलपणे आढळणारा गट आहे. या गटातील प्राणी उत्तर मायोसीन काळात अवतरले व रीसेंट काळात त्यांची कमाल भरभराट झाली. त्यांचा विकास मुख्यत्वे पूर्व गोलार्धात झाला. यांचे बहुसंख्य वंश व आफ्रिकेतील काही वंश दक्षिण आशियात व दक्षिण यूरोपात निर्माण झाले. त्यांच्या कवटीची हाडे भरीव वा हवेच्या पोकळ्यांनी युक्त असून नेत्रकोटराच्या पश्च भागात उपांग शाखा नर व मादीत आणि काही बाबतींत फक्त नरात असतात. यांच्या दंतविन्यासाचे सामान्य सूत्र ०.०.३.३/ ३.१.३.३ असून कृंतक दात व सुळे चमच्यासारखे असतात कपोल दात सेलोनोडोंट,ब्रॅकिओडोंट काही विशेषीकृत प्राण्यांत हिप्सोडोंट प्रकारचे असतात. दंतवल्काचा थर साधा क्वचित सुरकुत्या पडलेला असतो. उपदाढा आखूड व खालच्या दाढा अरुंद असतात. मुख्य पश्चपादास्थी (म्हणजे क्रमांक तीन व चार) एकत्र सांधल्या जाऊन नालास्थी बनलेली असते. ललाटीय प्रदेश सामान्यतः रुंद असून विशेषतः नेत्रकोटराच्या पश्च बाजूस तो रुंद असतो. त्यामुळे पार्श्वकपालास्थी पश्चकपालास्थीवर मागे ढकलल्या गेल्यासारख्या असतात. या बाबतीत बॉस हा वंश हे पराकाष्ठेचे उदाहरण आहे. त्यामध्ये जवळजवळ सर्व कवटी ललाटास्थीखाली असते व पार्श्वपालास्थी अल्पशीच दिसते. नेत्रकोटरांमध्ये अस्थिमय प्रवर्ध असून ते उभे व मागे वाकलेले असतात परंतु प्रथम प्राण्यांत हे प्रवर्ध मागे व कवटीच्या पश्च बाह्य कोपऱ्याच्या दिशेत बाहेरील बाजूस सरकलेले असतात उदा., बॉस.
सर्व्हिकॉर्नियांशी तुलना केल्यास कॅव्हीकॉर्निया हा प्रगतीशील गट असून कित्येक बाबतींत त्याचे विशेषीकरण झालेले असून अशा प्रकारे त्यात पुष्कळ विविधता आलेली आढळते. मात्र अशी विविधता सामान्य वा संबंधित वैशिष्ट्यांचे एकीकरण होऊनही आलेला असते. त्यामुळे सर्वसाधारण वैशिष्ट्यांच्या आधारे यांचे एकत्रित गट केल्यास त्यांची पुढील दोन उपकुले होतात : (१) ईगोडोंशिया व (२) बूडोंशिया यांचेही कित्येक उपगट पडतात.
ईगोडोंशियांची कवटी सर्वसामान्यपणे चांगली चापाकार असते. दाढा हिप्सोडोंट प्रकारच्या व शिंगे सामान्यतः बाजूने दबलेली व मागे वळलेली, क्वचित सरळ वा मळसूत्री असून ती नेहमी नेत्रकोटरांलगत वरच्या बाजूंवर आलेली असतात. या उपकुलाचे ५ गट असून त्यांपैकी निओट्रॅजिनी व गॅझेलिनी (पूर्वी हे अँटिलोपिनी या गटात एकत्रित केले होते) हे त्यांच्या हिप्सोडोंट प्रकारच्या दातांमुळे ओव्हिकॅप्रिनी गटाला अगदी जवळचे आहेत. या तीनही गटांचा उदय बहुधा उत्तर अमेरिकेतील अँटिलोकॅप्रिडी कुलापासून झाला असावा. निओट्रॅजिनीचे जीवाश्म केवळ आफ्रिकेतील रीसेंट कालीन खडकांत सापडतात. रुपिकॅप्रिनीचे जीवाश्म यूरोपातील प्लाइस्टोसीन कालीन खडकांत क्वचित आढळतात अन्यथा हा यूरोप व पश्चिम आशियातील रीसेंट कालीन गट आहे. गॅझेलीनी, ओव्हिकॅप्रिनी व ओव्हिबोव्हिनी यांचे प्लायोसीन काळापासूनचे जीवाश्म आढळतात व ते यूरोप, आशिया, आफ्रिका, उत्तर अमेरिका, ग्रीनलंड इ. भागात विस्तृतपणे पसरलेले आहेत. सध्या पाळल्या जात असलेल्या शेळ्या (कॅप्रा) व मेंढ्या (ओव्हिस) नवाश्मयुगात अवतरल्या होत्या.
बूडोंटिडी हा पूर्व गोलार्धातील गट असून तो बहुधा यूरोपमध्ये उत्तर (कदाचित पूर्व) मायोसीन काळामध्ये काहीशा अँटिलोपसारख्या वंशज असलेल्या व काहीसे खोलगट माथे असलेले हरिणासारखे दात असलेल्या सर्व्हीकॉर्नियांपासून निर्माण झाला असावा . हा प्रकार कमी अधिक प्रमाणात ट्रॅजिलॅफिनी व सेफॅलोफिनी गटांत टिकून राहिलेला आढळतो. ब्यूबॅलिनी, हिप्पोट्रॅजिनी व सर्व्हिकॅप्रिनी यांच्यातील दंतविन्यास पुष्कळसा बोव्हिनीसारखा असतो हे दात हिप्सोडोंट प्रकारचे असून त्यांच्या तळाशी स्तंभाकार रचना विकसित झालेली असते आणि कित्येकदा पुष्कळ संधानक असतो तसेच उपदाढा क्वचितच लहान झालेल्या आढळतात. ललाटास्थी मागील बाजूस पसरलेली असून तिने जवळजवळ सर्व कवटी झाकली गेलेली असते ही अवस्था बॉसमध्ये वैशिष्ट्यपूर्ण स्थितीत आढळते. शिंगे नुसती वाकडी व मागे वळलेली व कधीकधी मळसूत्रासारखी असतात. बोव्हिनीत शिंगे मागील बाजूला सरकलेली दिसतात आणि बॉस वंशात ती कवटीच्या पश्च-पार्श्व कोपऱ्याशी असतात. गव्याची शिंगे दंडगोलाकार असून ती वर व बाहेरील बाजूस वळलेली असतात. म्हशींचा ललाटीय भाग पुष्कळसा चापाकार असतो, तर इतरांत तो कमीअधिक प्रमाणात सपाट असतो.
स्यूडोट्रॅजिनीचे बहुतेक जीवाश्म यूरोपातील (शिवाय पर्शिया, चीन व उत्तर अमेरिका येथीलही) उत्तर मायोसीन ते पूर्व प्लायोसीन काळातील खडकांत सापडले आहेत. सर्व्हिकॅप्रिनी हा अगदी मर्यादित गट असून त्याचे जीवाश्म केवळ भारतातील प्लायोसीन व अल्जीरियातील प्लाइस्टोसीन खडकांत सापडले आहेत. ट्रॅजिलॅफिनीचे जीवाश्म उत्तर अमेरिका, यूरोप, उत्तर आफ्रिका व आशिया येथील प्लायोसीन ते प्लाइस्टोसीन कालीन खडकांत सापडले आहेत व हे प्राणी हल्ली आफ्रिकेत राहत आहेत.
गायी, म्हशी, गवे इ. बोव्हिनीमध्ये येतात व हे आर्टिओडॅक्टिलांपैकी सर्वांत जास्त उत्क्रांत झालेले प्राणी आहेत. ते प्लायोसीनमध्ये अवतरले व प्लाइस्टोसीनपासून ते विपुल आहेत. रानम्हशी हल्ली दक्षिण आशियात आढळतात. गव्याचे सर्वांत जुने जीवाश्म उत्तर प्लायोसीन कालीन असून ते उत्तर भारतात आढळतात ते यूरोप व उत्तर आशियात प्लाइस्टोसीनमध्ये विपुल होते. हल्लीचे यूरोपीय गवे लिथुॲनिया प्रदेशातील असून त्यांची अमेरिकन जाती उत्तर अमेरिकेत आहे. सध्या गाईबैल सर्व जगभर पसरलेले आहेत. त्यांचे सर्वांत जुने जीवाश्म उत्तर भारतातील शिवालिक संघातील खडकांत आढळले आहेत.
(८) नोटोअंग्युलेटा गण : शाकाहारींचा हा गण निर्वंश झालेला असून याचे जीवाश्म केवळ उत्तर अमेरिकेतील इओसीन ते प्लाइस्टोसीन कालीन खडकांत सापडले आहेत. मांसाहरी व रोडेंशिया यांच्याशी या गणाचे सर्वसाधारण साम्य असून या गणातील प्राणी खोकडापासून टॅपिरपर्यंतच्या आकारमानांचे होते. त्यांच्या कवटीचा माथा सपाट व रुंद होता आणि अनुकपाल रुंद होते अधोहनू उंच व पुष्कळदा भरीव असे. मुळात दंतविन्यास पूर्ण होता व सर्वांत पुढच्या कृंतक दातांचे आकारमान वाढण्याची प्रवृत्ती होती परंतु काही गटांत काही कृंतक लहान झालेले किंवा काही कृंतक व सुळेही नष्ट झालेले किंवा सुळे हत्तीच्या दातांसारखे झालेले दिसतात. कपोल दात ब्युनोडोंट प्रकारचे होते व विशेषीकृत प्राण्यांत ते हिप्सोडोंट प्रकारचे झालेले असत ते त्रिगुलिकायुक्त ते त्रिगुलिकायुक्त-त्रिज्याखंडी होते. यांचे अंग्युलेटाऐवजी पुष्कळसे कृंतक व क्रिओडोंट दात मांसाहारीप्रमाणे होते. बोटांच्या टोकांशी बोथट नख्या किंवा खूर असत. ते पादतलचारी ते अर्धपादतलचारी होते.
(९) सब-अंग्युलेटा गण : हे शाकाहारी प्राणी मोठ्या आकारमानाचे असून ते पादतलचारी ते अर्धपादतलचारी आहेत तर काही जलचर आहेत. आधीच्या प्राण्यांची कवटी बसकट लांबट होती व अनुकपाल रुंद होते. तसेच सममितार्धी कटक (मध्यभागी आडव्या दिशेत लांबट झालेला माथा) व अरुंद कवटी विकसित झालेली होती. मुळात दात ब्रॅकिओडोंट-ब्युनोडोंट प्रकारचे होते दाढा चतुर्गुलिकायुक्त असून दाताचे माथे कटकाने जोडलेले होते. खालच्या जबड्यात हे कटक कोरीसारखे असतात. उपदाढांचे दाढांत रुपांतर होण्याची प्रवृत्ती दिसते. कृंतक दातांच्या एका जोडीचा सामान्यतः हत्तीच्या मोठ्या दातांसारख्या दातांमध्ये म्हणजे हस्तिदंतामध्ये विकास झालेला आढळतो किंवा कृंतक ते सुळे ते उपदाढा असा त्यांच्या सामान्य स्वरूपात सावकाश होत गेलेला बदल आढळतो. जीवनप्रणालीनुसार पायांमध्ये विविध दिशांनी परिवर्तने झालेली आहेत.
सब-अंग्युलेटाचे पुढील चार उपगण केले जातात: (क) एंब्रिथोपोडा, (ख) हायरॅकॉयडिया, (ग) सायरेनिया, (ध) प्रोबॉसिडिया. हे उपगण ईजिप्तमध्ये उत्तर इओसीन काळात अवतरले. ते जवळजवळ त्या वेळच्या ब्युनोडोंट प्रकारच्या वंशजांपासून (बहुधा काँडिलार्थ्रापासून) आले असावेत, बाहेरून हे गट एकमेकांपासून काहीसे वेगळे दिसतात तरीही त्यांतील कवटीची व इतर संरचनांची साम्ये ही त्यांना एका गणात समाविष्ट करण्यास पुरेशी आहेत. अशा तऱ्हेने आधीच्या हायरॅकॉयडियांचा ब्युनोडोंट दंतविन्यास आणि आणि बाह्य वैशिष्ट्ये ही पुष्कळशी आधुनिक प्राण्यांसारखी आहेत परंतु मीरिकोथेरायडी हा गट आदिम प्रोबॉसिडिया व सायरेनिया यांना ब्युनोडोंट दंतविन्यास आणि कटी, स्कंधास्थी व भुजास्थी यांची रचना यांच्या बाबतींच जवळचा आहे. एंब्रिथोपोडांचे पाय प्रोबॉसिडियांप्रमाणे असतात, दंतविन्यास मात्र निराळा असतो.
सब-अंग्युलेटाचे पुढील चार उपगण केले जातात:(क)एंब्रिथोपोडा, (ख)हायरॅकॉयडिया (ग) सायरेनिया (घ)प्रोबॉसिडिया. हे उपगण ईजिप्तमध्ये उत्तर इओसीन काळात अवतरले. ते जवळजवळ त्यावेळच्या ब्युनोडोंट प्रकारच्या वंशजांपासून (बहुधा कॉंडीलॉर्थांपासून) आले असावेत, बाहेरून हे गट एकमेकांपासून काहीसे वेगळे दिसतात तरीही त्यातील कवटीची व इतर संरचनांची साम्ये ही त्यांना एका गणात समाविष्ट करण्यास पुरेशी आहेत. अशा तऱ्हेने आधीच्या हायरॅकॉयडियांचा ब्युनोडोंट दंतविन्यास आणि बाह्य वैशिष्टे ही पुष्कळशी आधुनिक प्राण्यासारखी आहेत परंतु मीरीकोथेरायडी हा गट आदिम प्रोबॉसिडिया व सायरेनिया यांचा ब्युनोडोंट दंतविन्यास आणि कटी, कंधास्थी व भूजास्थी यांची रचना यांच्या बाबतीत जवळचा आहे. एंब्रिथोपोडांचे पाय प्रोबॉसिडियांप्रमाणे असतात, दंतविन्यास मात्र निराळा असतो.
(क) एंब्रिथोपोडा उपगण : यातील प्राणी अगडबंब होते, त्यांचे पाय आखूड आणि मजबूत असून आदिम प्राण्यांच्या पायांना प्रोबॉसिडियांप्रमाणे पाच बोटे होती. यांची कवटी मोठी, मुस्कट आखूड व उंच आणि टिटॅनोथेरायडीप्रमाणे ह्यांच्या नासाप्रदेशावर शिंगे होती. दंतविन्यास परिपूर्ण होता छिन्नीसारखे कृंतक दात, सुळे व उपदाढा क्रमाने होत्या. दाढांना कोरीच्या आकाराचे कटक होते. पुढील पायांना पाच बोटे व त्यांवर प्रोबॉसिडियांसारखे खूर होते. मागील पाय अँब्लिपोडांसारखे असून रुंद बोटांना खूर होते. या गटातील आर्सिनोथेरायडी कालीन आर्सिनोथेरियम या एकाच वंशाचे ऑलिगोसीन कालीन जीवाश्म ईजिप्तमध्ये सापडले आहेत.
(ख) हायरॅकॉयडिया उपगण : हे प्राणी कृंतक प्राण्यांसारखे असून पादतलचारी ते अर्धपादतलचारी आहेत. त्यांच्या पुढील पायांना चार, तर मागील पायांना तीन बोटे असतात. खालचे कृंतक दात छिन्नीसारखे असून ते लहान झालेले असू शकतात. सुळे असतात किंवा लोप पावतात. उपदाढा कमीअधिक प्रमाणात दाढांसारख्या झालेल्या असून दाढा ब्युनोडोंट ते लोफोडोंट प्रकारच्या असतात. खालच्या जबड्याची वर जाणारी शाखा विशेष प्रकारे रुंदावलेली असते.
हायरॅसिडी कुलातील प्राणी पुष्कळसे कृंतक प्राण्यांसारखे असून ते लहान व चापट कवटी असलेले असतात. ते पादतलचारी असून त्यांच्या बोटांच्या टोकांवर पसरट नखे असतात. ते दक्षिण व पूर्व आफ्रिका, अरबस्तान व सिरिया या प्रदेशात राहत असून त्यांचे जीवाश्म आढळलेले नाहीत.
मायोहायरॅसिडी कुलाचे अपूर्णावस्थेतील जीवाश्म नैऋत्य व पूर्व आफ्रिकेतील मायोसीन कालीन खडकांत सापडले आहेत.
सॅगाथेरायडी कुलाचे जीवाश्म ईजिप्तमधील ऑलिगोसीन व प्लायोसीन कालीन खडकांत सापडले आहेत. त्यांचे आकारमान खोकडापासून टॅपिरपर्यंत एवढे असून त्यांचे दात ब्युनोडोंट ते लोफोडोंट प्रकारचे होते.
(ग) सायरेनिया उपगण : (समुद्र-गाय). हे शाकाहारी सस्तन प्राणी समुद्रकिनाऱ्यावर किंवा मोठ्या नद्यांच्या अधःप्रवाहात राहतात. हा सब-अंग्युलेटांचा दिसण्यात वैचित्र्यपूर्ण असा गट आहे. त्यांच्या शरीराची बाह्याकृती दंडगोलाकार दिसते व सामान्यतः ते भरभक्कम चणीचे असतात. हे बाहेरून पुष्कळसे सीटॅसियांसारखे (देवमाशांसारखे) दिसत असले, तरी अगडबंब शरीर, मोठी कवटी, दंतविन्यास व कटी यांबाबतींत त्यांचे अंग्युलेटांशी संरचनात्मक साम्य आढळते. यांचे दात एकवारदंती असून कपोल दात ब्युनोडेंट ते लोफोडोंट प्रकारचे असतात यांच्या वरच्या दाढा सहा गुलिकायुक्त व खालच्या दाढा चतुर्गुलिकायुक्त असून त्या पुष्कळशा डुक्कर व टॅपिर यांच्या दाढांप्रमाणे असतात. उपदाढा दाढांपेक्षा किंचित निराळ्या असतात सुळे बहुधा नसतात. जलचर जीवनप्रणालीस अनुरुप असे पायांचे वल्ह्यांसारख्या रचनेत परिवर्तन झालेले असते, त्यासाठी पायांची हाडे लवचिकपणे जोडलेली असून पायांना पाच बोटे (टोके) असतात. मागील पायांचा क्षय झालेला असतो. पुच्छपक्ष क्षितिजसमांतर असतो. सायरेनियांचे इओसीन काळापासूनचे जीवाश्म ईजिप्त, इटली, फ्रान्स व यूरोपातील इतर देश, दक्षिण अमेरिका, जपान यांसारख्या जगातील विविध भागांत सापडले आहेत. सध्या यांचे मेनॅटस व हेलिकोर हे दोनच वंश आफ्रिका, भारत आणि मध्य व दक्षिण अमेरिका यांच्या किनाऱ्यांवर आढळतात.
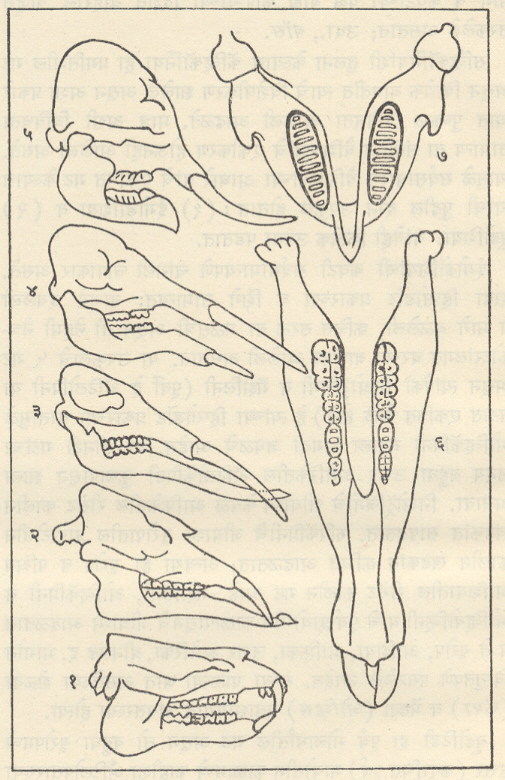
(घ) प्रोबॉसिडिया उपगण : (हत्ती). हे प्रचंड आकारमानाचे शाकाहारी अर्धपादतलचारी व सोंड असलेले प्राणी आहेत. त्यांची कवटी अजस्त्र असून तिच्यात मोठे वायुकोश असतात. प्राथमिक प्राण्यांचा दंतविन्यास परिपूर्ण होता. त्यांच्या दोन्ही जबड्यांत मोठ्या झालेला कृंतक दातांची जोडी होती पण नंतर खालच्या जबड्यातील ही जोडी नाहीशी झाली. आदिम प्राणी वगळता यांना सुळे नसतात. कपोल दात ब्युनो-लोफोडोंट ते लोफोडोंट प्रकारचे अथवा जवळजवळ असलेल्या आडव्या कटकांच्या मालिका व मधल्या गर्तांमध्ये (खाचांमध्ये) संधानक असलेले असे असतात. एलिफंटिडी कुलामध्ये कवटी व दंतविन्यास यांमध्ये आश्चर्यकारक परिवर्तने झालेली आहेत. पायांना पाच बोटे असून त्यांच्यात अल्पसाच बदल झालेला दिसतो. लांब हाडांमध्ये मज्जागुहिका (संयोजी ऊतकाने भरलेल्या मधल्या पोकळ्या) नसतात. ऊर्वस्थीला क्वचित तिसरे उरूकूट नसते, अंतर्जंघास्थी व बाह्य जंघास्थी तसेच प्रबाहू व प्रबाहु-अंतरास्थी या वेगवेगळ्या व बळकट असतात. बोटांना खुरांसारखी नखे असतात.
मोइरिथेरायडी कुल ईजिप्तमध्ये उत्तर इओसीन काळात अवतरले व ऑलिगोसीनच्या शेवटी निर्वंश झाले. त्यातील प्राणी टॅपिरएवढे होते. कवटीची रचना व दंतविन्यास यांवरून हे कुल सायरेनिया आणि प्रोबॉसिडिया यांच्यातील संक्रमणी अवस्था दर्शविते व या दोन्हींची समाईक पूर्वजपरंपरा दाखविते. त्यांच्या दातांचे सूत्र ३.१.३.३/२.०.३.३ असे होते दोन्ही जबड्यांतील कृंतक दातांची जोडी मोठी झालेली होती. व दाढांचे माथे कटकांनी जोडलेले असत.
बेरूथेरायडी कुलाचे जीवाश्म ईजिप्तमधील उत्तर इओसीन कालीन खडकांत आढळले आहेत. मात्र या कुलाचे वर्गीकरणातील स्थान अनिश्चित आहे. त्यांचे शरीर भरदार होते, दातांचे सर्वसाधारण सूत्र १.०.३.३/१.१.३.३ असे होते आणि त्यांचा खालचा जबडा पुष्कळसा डायनोथेरियमसारखा होता.
डायनोथेरायडी कुलातील प्राणी आफ्रिकेतील मायोसीन ते प्लाइस्टोसीन काळात, तर युरोप व आशियातील मायोसीन ते प्लायोसीन काळात होते. भारतामध्ये शिवालिक टेकड्यांतील पूर्व प्लायोसीन खडकांत डायनोथेरियमचे जीवाश्म आढळले आहेत. यांच्या खालच्या जबड्याच्या प्रतरसंधी खाली तर दात मागे वळलेले होते. कवटी उंच व भालप्रदेश रुंद होता. वरच्या दाढांवर तीन वा दोन कटक असत आणि खालच्या दाढांची रुंदी लांबीपेक्षा जास्त होती. नासाद्वारे (नाकपुड्या) बरीच वर उचलून ती लांब सोंडेची सूचक होती. यांच्या काही जाती हत्तीपेक्षा मोठ्या होत्या.
कवटी व दंतविन्यास यांच्या बाबतीत आश्चर्यकारक परिवर्तनांसाठी एलिफंटिडी हे कुल खासकरून प्रसिद्ध आहे. यात जबडे आखूड होण्याची प्रवृत्ती आढळते. भालप्रदेश उभट होत जाऊन कवटी उंच होत होती. वरच्या ओठाचे रुपांतर सोंडेत झाले व जसजशी शरीराची उंची वाढत गेली तसतशी सोंड लांब होत गेली. मोठे डोके व सोंड यांचा भार तोलून धरण्यासाठी पश्चकपालाचे क्षेत्र पुष्कळच वाढले. सोंडेचा विकास होताना मधल्या कृंतक दाताचे दमन झाले. दीर्घस्थायी गोर्द-गुहा असलेल्या कृंतक दातांच्या जोडीचे आपण ज्यांना हत्तीचे सुळे म्हणतो त्या मोठ्या दातांत म्हणजे हस्तिदंतांत परिवर्तन झाले आणि खालच्या जबड्यातील हे दात प्रगत जातीत नष्ट झाले. आदिम प्राणी वगळता इतरांना उपदाढा नसतात. आधीच्या प्राण्यांत कायमचे दात खालून वर, तर प्रगत प्राण्यांत मागून पुढे ढकलले जाऊन दुधाच्या कपोल दातांची जागा त्यांनी घेतली. हिप्सोडोंट प्रकारच्या मोठ्या दाढांसाठी पुरेशी जागा व्हावी म्हणून उत्तर हन्वस्थी (वरच्या जबड्याचे हाड) मोठी झाली आणि कटकांची संख्या पॅलीओलोक्झोडोंट अवस्थेतील ३ पासून एलिफसमधील जटिल दाढांवरील २७ कटकांपर्यंत वाढली. दंतविन्यासाच्या स्वरूपानुसार कित्येक वंश व उपवंश पाडले जातात. हे वंश व उपवंश एकमेकांपासून वेगळे ओळखणे नेहमीच सोपे नसते व काही तर प्रादेशिक परिवर्त (विशेष प्रकार) आहेत. म्हणून येथे अधिक नमुनेदार वंश (किंवा अगदी निकटचे नातेसंबंध असलेल्या वंशांचे गट) विचारात घेतले आहेत.
पॅलीओमॅस्टोडॉनची सममितार्धी कटक असलेली कवटी मध्यम उंचीची होती प्रत्येक जबड्यात दोन आखूड हस्तिदंत होते व खालचे दात चमच्याच्या आकाराचे होते. सर्व दाढा सलग सरळ रेषेत असून दंतमूलांच्या तीन जोड्या कटकांनी जोडलेल्या असत. ईजिप्तमधील ऑलिगोसीन कालीन खडकांमध्ये या वंशाचे जीवाश्म आढळले आहेत.
टेट्राबेलोडॉन वंशात मस्तिष्क-गुहा कमी उंच होती व ती एलिफसमध्याप्रमाणे पुष्कळच उंच होत गेली परंतु भालप्रदेश कमी उभट होता. अधोहनू लांब होती. उपदाढांना चांगले वाढलेले ३ ते ५ कटक असून त्यांद्वारे दंतमूलांच्या जोड्या सांधलेल्या असत. खालच्या दाढा त्यामानाने अरूंद होत्या. दातांच्या माथ्यावर दंतवल्क होते व त्यांच्या मध्यभागी लांबीच्या दिशेत खोबण होती. वरचे हस्तिदंत खालच्यापेक्षा लांब होते. उत्तर अमेरिका, यूरोप व पाकिस्तान येथील मायोसीन व प्लायोसीन कालीन खडकांत या वंशाचे जीवाश्म सापडले आहेत. भारतात सौराष्ट्राच्या दक्षिणेचे पिराम बेट व शिवालिक टेकड्या येथील याच काळातील खडकांत याचे जीवाश्म आढळले आहेत.
मॅस्टोडॉन वंशात अधोहनू एलिफसप्रमाणे आखूड झालेली होती. दाढा ब्युनो-लोफोडोंट ते लोफोडोंट प्रकारच्या होत्या. टेट्राबेलोडॉनपेक्षा याची अधोहनू आखूड होती. यूरोप, दोन्ही अमेरिका, पर्शिया, पाकिस्तान, चीन व जपान येथील प्लायोसीन कालीन तसेच उत्तर अमेरिकेतील प्लाइस्टोसीन कालीन खडकांत या वंशाचे जीवाश्म आढळले आहेत. भारतात याचे प्लायोसीन कालीन जीवाश्म पिराम बेटावर व शिवालिक टेकड्यांत सापडले आहेत.
स्टेगॉडॉन हा मॅस्टोडॉन व एलिंफस (खरे हत्ती) या गटांतील संक्रमणी गट मानतात . मॅस्टोडॉनमध्ये खालचे हस्तिदंत नसत. दाढांवर बहुपिंडिकीय (अनेक गाठी असलेले) ६ ते १२ कटक असून त्यांचे माथे किंचित बहिर्गोल असतात व गर्त संधानकाने भरलेले असतात. दक्षिण व पूर्व आशिया व जावा येथील मध्य आणि उत्तर प्लायोसीन कालीन खडकांत स्टेगोडॉनचे जीवाश्म आढळले आहेत. भारतामध्ये शिवालिक टेकड्या व जम्मू येथील मध्य व उत्तर प्लायोसीन कालीन खडकांत आणि नर्मदा व गंगा खोऱ्यातील प्लाइस्टोसीन कालीन गाळात स्टेगोडॉनचे जीवाश्म सापडले आहेत.
एलिफस वंशांत दातांचे सूत्र १.०.०.३/०.०.०.३ असे आहे. त्यांच्या दाढा हिप्सोडोंट प्रकारच्या असून त्यांच्यावर जवळजवळ असलेले असे अनेक (२७ पर्यंत) चापट कटक असतात. उदा., एलिफस प्रायमिजिनियस व ए. मॅक्झिमस यांच्यात २७ कटक आहेत. अशा प्रकारे या दातांच्या माथ्यावर संधानक-दंतवल्क व दंतिन यांचे एकाआड एक असे पट्टे दिसतात. या वंशाचा (किंवा अगदी निकटचे नातेसंबंध असलेल्या वंशाच्या गटाचा) प्रसार प्लायोसीन व प्लाइस्टोसीन काळांत यूरोप-उत्तर आफ्रिका, दोन्ही अमेरिका आणि आशिया या भागांत विस्तृतपणे झाला होता. हा वंश प्रथम भारतात अवतरला. याचे जीवाश्म शिवालिक टेकड्यांतील प्लायोसीन कालीन खडकांत, तसेच नर्मदेच्या प्लाइस्टोसीन कालीन गाळात व जम्मूतील कारेवा खडकांत सापडले आहेत. या वंशाचे प्राणी हल्ली भारत, आफ्रिका, ब्रह्मदेश व आग्नेय आशिया येथे प्रादेशिक परिवर्त म्हणून आढळतात. यातील ए. मेरियायोनॉलिस व ए. अँटिक्कस यांसारख्या जातींतील प्राण्यांची उंची ४ मी. होती. जीवाश्मांपैकी ⇨ मॅमथ (ए. प्रायमिजिनियस) या ४ मी. लांबीचे हस्तिदंत असलेल्या जातीचा भौगोलिक प्रसार बहुधा जास्तीत जास्त झालेला होता ही जाती फिनलंड व स्कँडिनेव्हिया सोडून सर्व यूरोप, उत्तर अमेरिका, उत्तर आफ्रिका, कॅस्पियन समुद्र व बैकल सरोवरापर्यंतचा उत्तर आशिया, चीन व जपान इ. भागांत होती.
(१०) नरवानर गण [प्रायमेट्स ⟶ नरवानर गण]. मानव, कपी, माकड, लेमूर, वृक्ष-चिचुंदरी हे नरवानर गणातील प्राणी असून ते सर्वभक्षी असतात आणि शाकाहारी व फलभक्षी होण्याकडे त्यांची प्रवृत्ती दिसते. मऊसर आहारामुळे त्यांच्या दातांचे इतर सस्तन प्राण्यांच्या मानाने विशेषीकरण अतिशय कमी झालेले आहे. तथापि काही प्रकारच्या दातांची संख्या कमी आढळते. अशा प्रकारे मंगोलियातील क्रिटेशस कालीन खडकांत आढळलेल्या ट्युपाइऑइडियामधील ॲनेगेले या नरवानरांच्या सर्वांत आदिम गटामध्ये दातांचा पूर्ण पूरक संच होता व त्यांना पुढील सूत्रात दर्शविल्याप्रमाणे एकूण ४४ दात होते
३.१.४.३/३.१.४.३ यूरोप व उत्तर अमेरिकेत आढळलेला पॅलिओसीन कालीन प्लेसिॲडेपिस हा दुसरा अगदी आदिम गट जी. जी. सिम्पसन यांच्या मते ट्युपाइऑइडियाशी तसेच लेमुरॉडियाशी संबंधित आहे. ट्युपाइऑइडियामधील आणि झाडांवर राहणाऱ्या हल्लीच्या वृक्ष-चिचुंदरीमधील दातांचे सूत्र २. १.३.३/२.१.३.३ असे, तर सध्याच्या लेमूरच्या दातांचे सूत्र २.१.२.३/२.१.२.३ असे, आहे. अशा प्रकारे कृंतक दातांची संख्या कमी होऊन अगदी आदिम नरवानरांतही ती दोन झालेली आढळते व बहुतेक नरवानरांत सामान्यपणे दोनच कृंतक दात असतात. तथापि काहींमध्ये (उदा., आय-आय प्राण्यामध्ये) दोन्ही जबड्यांतील एकेक कृंतक दात कमी झालेला असतो, तर टार्सिअरमध्ये खालच्या जबड्यात एकक कृंतक असतो फ्रान्समधील इओसीन कालीन नेक्रोलेमूर व उत्तर अमेरिकेतील इओसीन कालीन टेटोनियस यांना खालचे कृंतक नव्हतेच. वरील प्लेसिॲडेपिसमध्ये व हल्लीच्या आय-आयमध्येही कृंतक मोठे झालेले आढळतात परंतु एकंदरीत उच्च नरवानरांमधील कृंतक दातांत थोडेच बदल झालेले आहेत. सुळ्यांमध्येही थोडेच बदल झालेले दिसतात. सामान्यपणे ते कृंतकपेक्षा किंचित उंच असतात पण पूर्व गोलार्धातील माकडे व काही कपी यांचे सुळे अधिक लांब असून बहुधा सुळा व शेवटच्या कृंतक यांच्यात दंतावकाश असतो.
ॲनॅगेलेसारख्या आदिम जीवाश्मांत चार उपदाढा होत्या परंतु त्यांच्या हल्लीच्या प्रातिनिधिक प्राण्यांमध्ये उपदाढा तीनपेक्षा जास्त नसतात, तर उच्चतर प्राण्यांमध्ये दोनच उपदाढा असतात. विशेषकरून त्यांना दोन दंताग्रे असू त्यांचे दाढांत रुपांतर होण्याची प्रवृत्ती दिसत नाही. ज्या दोन उपदाढा नाहीशा झाल्या त्या अग्र बाजूच्या होत्या. आय-आयमध्ये फक्त वरची एकच उपदाढ असते व खालच्या उपदाढा नसतात. तथापि निर्वंश झालेल्या इओसीन कालीन टार्सिऑइडियांच्या दाढांमध्ये काही रचनात्मक परिवर्तने झालेली आढळतात. आदिम नरवानरांच्या दाढांची रचना आदिम सस्तन प्राण्यांमध्ये त्रिगुलिकायुक्त होती परंतु गुलिका बोथट किंवा अग्रवर्धी (टोकदार) असून चवथे दंताग्र निर्माण होण्याच्या प्रवृत्तीमुळे त्यांना ब्युनोडोंट स्वरूप प्राप्त होते. पूर्व गोलार्धातील माकडांमध्ये ही दंताग्रे आडव्या कटकाने जोडलेली असतात. मायोसीन कालीन प्रोकॉन्सूलसारख्या काहींमध्ये पाचवेही दंताग्र असते. कपींचे दात मूलतः चतुर्गुलिकायुक्त असले, तरी पुष्कळांमध्ये पाचवी गुलिकाही आढळते. मऊसर आहारामुळे त्यांचे दात ब्रॅकिओडोंट प्रकारचेच राहिले. लांब दाढा असलेला बॅबून हा अपवाद सोडल्यास सामान्यतः दातांची ओळ (दंतपंक्ती) आखूड होण्याच्या प्रवृत्तीमुळे त्याचे मुस्कट आखूड होत गेलेले दिसते.
आधीचे आदिम नरवानर व त्यांचे हल्लीचे वंशज, तसेच ज्याचा क्रमविकास कमी झाला आहे असे गट यांच्यातील प्राण्यांची कवटी लांबट व बुटकी मुस्कट पुढे आलेले असते परंतु क्रमविकास होताना गोलसर माथा असलेली कवटी उभट होत गेली व तिचे आकारमान वाढत गेले. यामुळे अंशत पुढे आलेली चेहेरेपट्टी उभट होत गेली व शेवटी ती मानवाप्रमाणे उभट झाली. दातांच्या ओळी आखूड होतांना मुस्कट आखूड होत गेल्यानेही चेहेरेपट्टी उभट होत जाण्यास मदत झाली.
माकड, कपी व मानव यांच्यातील ढुंगणावर बसण्याच्या आणि ताठ उभे राहून चालण्याच्या सवयीमुळे अंशतः तसेच डोक्याच्या मागील वाढीमुळे पश्चकपालास्थि-कंद मागील बाजूला जाऊन कवटीच्या खाली सरकला आहे.
कवटीतील दुसरा ठळक बदल म्हणजे लेमूर व कपी यांसारख्या आदिम प्राण्यांत असलेले सममितार्धी व पश्चकपालास्थी कटक मानवात नाहीसे झाले, हा होय.
नेत्रकोटरे पुढील बाजूस वळविली गेली व त्रिमितीय दृष्टी विकसित झाली. कारण अशी दृष्टी येणे वृक्षवासी व वनात राहण्याच्या जीवन प्रणालीत अंतराचा स्पष्ट व अचूक अंदाज येण्यासाठी अगदी आवश्यक आहे.
वृक्षवासी जीवनप्रणालीमुळे कवटीच्या मागील सांगाड्याचे स्वरूप आदिम सस्तन प्राण्यांच्या सांगाड्याप्रमाणे लवचिक राहिले. झाडांना लोंबकळपणे व झाडांवरून उड्या मारणे यांस साहाय्यभूत व्हावे म्हणून वक्षीय कमानीचा महत्त्वाचा भाग म्हणून जत्रू (गळपट्टीचे हाड) टिकून राहिले. चालण्याची पादतलचारी पद्धती व पाच बोटे असलेले पाय ही आदिम वैशिष्ट्ये असून ती टिकून राहिली आहेत. चतुष्पादांचे पुढचे पाय व द्विपादांचे हात हे परिग्राही (धरून व पकडून ठेवू शकणारे) अवयव असून हाताचा आंगठा व पायाचा आंगठा (विशेषतः पायाचा आंगठा) हे कमीअधिक प्रमाणात संमुख स्वरुपांचे (इतर बोटांच्या समोर आणता येतील असे) असल्याने वस्तू पकडण्यास त्यांची मदत होते. लेमूर व मार्मोसेट हे वगळता बोटांच्या टोकांचे संरक्षण व्हावे म्हणून नख्यांच्या जागी नखे आलेली आहेत. अंगस्थिती (बसण्याची ढब) सामान्यपणे चतुष्पादांप्रमाणे असून ती बदलून द्विपादांप्रमाणे व विशेषतः मानवात ताठ उभी अशी झाली.
नरवानर गमाने कार्ल फोन त्सिटेल व इतरांनी (क) प्रोसिमिआय, (ख) अँथ्रॉपॉइडिया व (ग) बायमॅना असे तीन तर जी. जी. सिम्पसन व इतरांनी याचे (क) प्रोसिमिआय व (ख) अँथ्रॉपॉइडिया असे दोन उपगण केले आहेत. मानव व त्याचे निकटचे नातेवाईक म्हणजे मानवसदृश (अँथ्रॉपॉइड) कपी हे इतर मानवसदृश प्राण्यांपेक्षा किती प्रमाणात वेगळे गणले जावे, याला देण्यात आलेल्या महत्वानुसार वरील दोन प्रकारे नरवानरांचे उपगण करण्यात आलेले आहेत.
किटकभक्षी प्राण्यापासून नरवानरांसह अपरास्तनी प्राण्यांचे पुष्कळ गट उदयास आले. मंगोलियात आढळलेला उत्तर क्रिटेशन कालीन डेल्टाथेरिडियम हा कीटकभक्षींचा सर्वांत जुना वंश आहे. यावरून अपरास्तनी प्राणी क्रिटेशसच्या अखेरीस किंवा पॅलिओसीनच्या प्रारंभी उदय पावले असणे शक्य आहे परंतु ते अवतरल्यानंतरच्या सु. ६ कोटी वर्षांच्या काळातील नरवानरांच्या जीवाश्मांच्या नोंदी इतर सस्तन प्राण्यांच्या मानाने अपूर्ण व तुरळक आहेत. नरवानरांच्या वृक्षवासी व वनात राहण्याच्या जीवनप्रणालीमुळे बहुधा असे घडले असावे, कारण ही परिस्थिती जीवावशेष गाडले जाण्यास व ते टिकून राहण्यास अनुकूल नाही.
(क) प्रोसिमिआय उपगण : नरवानरांच्या सर्वांत आदिम अशा ट्युपाइऑइडिया गटातील ॲनॅगेले वंशाचे जीवाश्म मंगोलियातील ऑलिगोसीन कालीन खडकांत आढळले आहेत परंतु तदनंतरच्या काळातील त्यांचे जीवाश्म जवळजवळ आढळत नाहीत. त्यांचे हल्लीचे वंशज म्हणजे लहान खारीसारख्या वृक्ष-चिचुंदऱ्या या असून त्या आग्नेय आशिया व ईस्ट इंडीज या प्रदेशात राहत आहेत.
लेमुरॉइडिया या लहान गटातील प्राणी कोल्ह्याप्रमाणे दिसतात व ते केवळ मॅलॅगॅसी व त्या जवळील कोमोरो बेटांवरच आहेत. त्यांचा क्रमविकास थोडाच झाला आहे. पॅलिओसीन कालीन प्लेसिॲडेपिस तसेच इओसीन कालीन ॲडेपिस व नेक्रोलेमूर या सर्वांत आधीच्या लेमुरॉइडियांमध्ये आढळणाऱ्या आदिम वैशिष्ट्यांचा मागोवा त्यांच्या तृतीय कालीन व हल्लीच्या वंशजांमध्ये घेता येतो. मॅलॅगॅसीत आढळलेल्या आधीच्या प्लाइस्टोसीन काळातील मेगॅलॅडेपिसची (मे. इन्सिग्निस ही याची एक जाती आहे) कवटी ३० सेमी. लांब असून तो नरवानरांपैकी सर्वांत मोठा प्राणी होता. त्यांचा दंतविन्यास, कवटीची वैशिष्ट्ये व शेपटी नसणे यांवरून हा प्राणी लेमुरॉडियांची एक अपवादात्मक वेगळी शाखा होती असे दिसते. आधुनिक इंड्रिस ही दुसरी अपवादात्मक वेगळी शाखा असून या शाखेचे प्लाइस्टोसीन कालीन जीवाश्म मॅलॅगॅसीत आढळले आहेत. डॉबेटोमिया किंवा केरोमिस म्हणजे आय-आय ही बहुधा लेमुरॉडियांची आणखी एक अपवादात्मक वेगळी शाखा असून तिचे माकडांशी बाह्यतः साम्य आहे. या शाखेचे प्लाइस्टोसीन काळातील जीवाश्म मॅलॅगॅसीत आढळले आहेत.
लोरेसिडी कुलातील प्राणी हे लेमूरांपेक्षा किंचित प्रगत आहेत. आशियाच्या उष्ण कटिबंधीय भागातील लोरिस (स्लो लेमूर) आणि आफ्रिकेच्या उष्ण कटिबंधीय भागातील गॅलॅगो (बुश बेबी) व पेरोडिसीयस (पोट्टो) हे यांचे हल्लीचे प्रतिनिधि आहेत. या गटाच्या जीवाश्मांच्या नोंदीही अल्प असून याचे पूर्व आफ्रिकेत मायोसीन कालीन प्रोगॅलॅगो व हिमाचल प्रदेशात प्लायोसीन (मध्य शिवालिक) कालीन इंद्रालोरिस या वंशांचे जीवाश्म सापडले आहेत.
सुमात्रा ते फिलपीन्स या भागात मोठ्या डोळ्यांचे टार्सिअस वंशाचे प्राणी राहतात. पुष्कळ आखूड झालेला नासाप्रदेश व पुढे रोखलेली मोठी नेत्रकोटरे यांच्यामुळे हे प्राणी पुष्कळसे माकडांसारखे दिसतात. टार्सिइडी कुलाचे थोडेच जीवाश्म उत्तर अमेरिका, चीन व युरोप येथील इओसीन व ऑलिगोसीन कालीन खडकांत आढळले आहेत.
(ख) अँथ्रॉपॉइडिया उपगण : प्लॅटिऱ्हिनी श्रेणीतील म्हणजे रुंदनासिका (नकट्या नाकाच्या) माकडांच्या नाकपुड्या काहीशा अलग व बाजूला फेंदारलेल्या असतात. पुष्कळांमध्ये शेपटी परिग्राही असते. प्रोसिमिआय प्राण्यांपेक्षा याची मस्तिष्क-गुहा मोठी असते. पायाचे आंगठे संमुख स्वरूपाचे तर हातांचे आंगठे केवळ अंशतः संमुख असतात. सेबिडी कुलातील प्राण्यांमध्ये तीन दाढा टिकून राहिलेल्या असतात (उदा., प्रोसिमिआय प्राणी) व सर्व बोटांना नखे असतात. या माकडांचे इओसीन काळापासूनचे जीवाश्म आढळतात परंतु मायोसीनपासून त्यांच्यात अल्पसाच बदल झालेला दिसतो.
कॅलिथ्रिसिडी कुलातील माकडे सर्वांत लहान असून त्यांना दोनच दाढा असतात फक्त पायांच्या आंगठ्यांवर नखे असून इतर बोटांना नख्या असतात. ही दोन कुले बहुधा ऑलिगोसीन काळात स्पष्टपणे स्वतंत्र झाली असावीत.
कॅटॅऱ्हिनी श्रेणीतील माकडे व त्यांचे जीवाश्म केवळ पूर्व गोलार्धात आढळतात. ही माकडे सर्कोपिथेसिडी कुलातील असून त्यांच्या नाकपुड्या खालील दिशेत तिरप्या रोखलेल्या असतात म्हणून त्यांना अधोनासिका माकडे म्हणतात. यांच्या हातापायांचे आंगठे संमुख असतात. दंतविन्यास २.१.२. ३/२.१.२.३ असा असतो आणि कपी व मानवातही तो असाच असतो. या कुलात पुष्कळ विविधता आलेली असून याचा विस्तृत प्रसार झाला आहे. या कुलाची दोन उपकुले आहेत. सर्कोपिथेसिनी उपकुलात आफ्रिकेतील बहुतेक सर्व अधोनासिका माकडे, उत्तर भारतातील बंदर व आग्नेय आशिया, सेलेबिस, जपान इ. प्रदेशांतील मॅकाकस माकडे येतात. कोलोबिडी उपकुलात आफ्रिकेतील कोलोबस, भारत व श्रीलंका येथील सेम्नोपिथेकस (पवित्र माकडे वा हनुमान), वानर (लंगूर) आणि त्यांच्याशी संबंधित प्रकार, तिबेटातील बसक्या नाकाची माकडे, मलेशिया व ईस्ट इंडीज येथील प्रेसबिटिस आणि बोर्निओतील नासावानर (लांब नाकचे वानर) ही येतात.
प्लायोसीनच्या आधीच्या काळातील सर्कोपिथेसिडी कुलातील माकडे, टार्सिऑइड प्राणी, लेमूर इत्यादींमधील नातेसंबंध जीवाश्मांवरून स्पष्ट झालेले नाहीत.
भारतामध्ये उत्तर शिवालिक (उत्तर प्लायोसीन) खडकांत आणि मद्रास व आसाम येथील पूर्व रीसेंट कालीन गाळात मॅकाकचे जीवाश्म सापडले आहेत. शिवालिक व सिमला टेकड्या, तसेच कर्नुल गुहा या प्रदेशांतील प्लाइस्टोसीन कालीन निक्षेपांत सेम्नोपिथेकस व पॅपिओ यांचे अवशेष सापडले आहेत.
होमिनॉयडिया अधिकुल : चालण्याची ढब, शेपटीचा अभाव व कमी विशेषीकृत दाढा या पाँजिडी कुलातील मानवसदृश कपींच्या वैशिष्ट्यांमुळे ती सर्कोपिथिसिडी कुलातील कपींपासून आणि माकडांपासून वेगळी ओळखू येतात परंतु त्यांच्यात काही संक्रमणी रचना आढळतात. अशा प्रकारे पाँजिडी कुलातील आधीच्या प्राण्यांचा (उदा., ब्रह्मदेशात आढळलेल्या उत्तर इओसीन कालीन अँफिपिथेकस व पोंडाँगिया या प्राण्यांचा) दंतविन्यास हा गिबन (हायलोबेटिनी) व सर्कोपिथिसिडींपैकी नेक्रोलेमूरिनी यांच्या दंतविन्यासांच्या मधला (दरम्यानचा) आहे. याचप्रमाणे ऑलिगोसीन-मायोसीन कालीन प्रॉप्लिओपिथेकसचा दंतविन्यास इओसीन कालीन टार्सिऑइड प्राण्यांच्या दंतविन्यासांसारखा होता.
पाँजिडी कुलातील प्राण्यांच्या प्रसार मायोसीन काळात पूर्व आफ्रिका ते युरोप व दक्षिण आशिया असा झाला, हे मानवसदृश प्राण्यांच्या विकासाचे व प्रसाराचे ठळक वैशिष्ट्य आहे. बहुधा ईजिप्तमधील ऑलिगोसीन कालीन प्रॉप्लिओपिथेकसपासून मायोसीन काळात मोठ्या आकारमानाच्या शिवपिथेकस केन्यामध्ये उदयास आला. बहुधा नंतर शिवपिथेकसपासून पूर्व प्लाइस्टोसीन काळात सिमला टेकड्यांत पोंगो (ओरँगउटान) उदयास आला व चीनमधील प्लाइस्टोसीनच्या आधीच्या काळातील जायगँटोपिथेकस ही बहुधा त्यांची शाखा आहे. केन्यातील पूर्व मायोसीन काळातील प्रोकॉन्सूलमध्ये लहान मस्तिष्क-गुहेसारखी आदिम प्रकारची वैशिष्ट्ये होती व बहुतकरून त्यापासून ड्रायोपिथेकस व ड्रायोपिथेकसच्या एका शाखेपासून अँथ्रॉपोपिथेकस (चिंपँझी) व दुसरीपासून गोरिला यांचा उदय झाला. भारतीय प्रदेश प्लाइस्टोसीन काळात मानवसदृश प्राण्यांच्या विकासाचे केंद्र होते. येथे शिवपिथेकस, ड्रायोपिथेकस, ब्रह्मपिथेकस, रामपिथेकस इ. जीवाश्म आढळतात. सर्वांत आधीची अशी मानवाची वैशिष्ट्ये (विशेषतः लहान सुळे, दाढांची साधी रचना व सपाट झीज आणि कमानदार टाळू) रामपिथेकसमध्ये आढळतात व बहुधा त्याच्यापासून होमिनिडी कुल निर्माण झाले असावे.
मायोसीन, प्लायोसीन व प्लाइस्टोसीन काळात पाँजिडी कुलाचा जलदपणे विकास व प्रसार झाला. त्यांचे हल्लीचे वंशज म्हणजे गिबन हे आसाम, पश्चिम युनान, ब्रह्मदेश, मलेशिया ते वोर्निओ या प्रदेशांत आढळतात. ओरँगउटान (पोंगो) सुमात्रा व बोर्निओ या भागांत तर चिंपँझी (अँथ्रॉपोपिथेकस) आफ्रिकेच्या उष्ण कटिबंधीय प्रदेशात आणि गोरिला आफ्रिकेच्या विषुववृत्तीय प्रदेशात राहत आहेत. ओरँगउटान वृक्षवासी तर गोरिला चिंपँझी अंशतः वृक्षवासी व अंशतः भूचर आहेत. सर्कोपिथेसिडी कुलाच्या मानाने पाँजिडी कुलातील आदिम प्राण्यांपासून हल्लीच्या प्राण्यांपर्यंत मस्तष्क-गुहेचे आकारमान थोडेच वाढले आहे.नरवानरांपैकी होमिनिडी कुलातील प्राण्यांचा क्रमविकास सर्वाधिक झालेला आहे. या कुलात मानव (होमो) व त्याला अगदी जवळचे असे प्राणी येतात. पाँजिडी व होमिनिडी कुलांच्या दरम्यानचे संक्रमणी प्राणीही आहेत. होमिनिडी कुलातील अधिक प्रगत (व त्यामुळे प्रातिनिधिक) प्राण्यांच्या पुढील वैशिष्ट्यांमुळे ते पाँजिडीतील प्राण्यांपासून वेगळे ओळखू येतात होमिनिडीतील प्रगत प्राण्यांच्या मस्तिष्कगुहेचे आकारमान पुष्कळ जास्त (पाँजिडी कुलातील आधुनिक प्राण्यांच्या जवळजवळ दुप्पट) असते त्यांचे कपाळ उंच, चेहेरेपट्टी जवळजवळ उभट व हनुवटी चांगली वाढलेली असते, पश्चकपालास्थी व सममितार्धी कटक आणि नेत्रकोटराचे उंचवटे नसतात, उत्तर हन्वस्थीच्या अग्र बाजूंचे हाडही नसते लहान झालेले सुळे व दंतावकाश नाहीसे झाल्याने दातांची ओळ आखूड व रूंद गोलाकार झालेली असते पायांची बोटे आखूड झालेली असून पायाचा आंगठा संमुख नसतो चालण्याची ढब ताठ उभी असते आणि हात पायांपेक्षा आखूड असतात व हालचाल दोन पायांद्वारे होते.
होमिनिडी कुलाचे जीवाश्म जवळजवळ फक्त प्लाइस्टोसीन काळातच आढळतात.
प्लाइस्टोसीन काळातील हिमयुगाच्या चार टप्प्यांत मानव-पूर्व व आधीचे मानव वंश नष्ट झाले व या दुःसह जलवायुमानीय बदलांना केवळ आधुनिक मानवच म्हणजे होमोसें पिएन्स (प्रगल्भ किंवा बुद्धिप्रधान मानव) यशस्वीपणे तोंड देऊन टिकून राहिला. प्लाइस्टोसीन मधील तात्पुरत्या कमी दुःसह म्हणजे काहीशा उबदार जलवायुमानाच्या काळात साचलेल्या अंतराहिमानीय (म्हणजे शीत जलवायुमानाच्या दोन काळांमधील उबदार जलवायुमानाच्या काळातील) निक्षेपांमध्ये आधुनिक मानवाचे जीवाश्म सापडतात. बहुधा दात, कमी अधिक प्रमाणात भंगलेल्या कवट्या व हातापायांची हाडे जीवाश्मरूपात आढळतात. वाजवी प्रमाणात चांगले टिकून राहिलेले व जवळजवळ पूर्ण असे थोडेच सांगाडे आढळले आहेत. १९२० च्या सुमारापासून मानवाचे जीवाश्म बऱ्याच प्रमाणात उपलब्ध झाले आहेत आणि त्यांच्यावरून पुष्कळ वंश व जाती प्रस्थापित करण्यात आल्या आहेत. येथे त्यांचे केवळ प्रमुख गट विचारात घेतले आहेत. [⟶ मानवप्राणि].
ऑस्टॅलोपिथेकस गट : झिंजांथ्रोपस हा पूर्व प्लायोसीन कालीन जीवाश्म ट्रान्सव्हालमध्ये सापडला असून प्लेसिअँथ्रोपस, पॅरँथ्रोपस व टेलँथ्रोपस हे त्याचे नमुने आहेत तसेच होमो हॅबिलिस हीही ट्रान्व्हालमधील जाती आहे. यांचे वर्णन त्यांच्या कवट्या व मानवासारखे दात त्यांच्यावरून केले आहे व ते अपमानवी दर्जा असलेले ऑस्टॅलोपिथेकसचे केवळ परिवर्त मानले जातात.
पिथेकँथ्रोपस इरेक्टस गट : चीनमधील सायनँथ्रोपस पीकिनेन्सिस (पीकिंग मानव) ही जाती जावाच्या पहिल्या अंतराहिमानीय निक्षेपातील पिथेकँथ्रोपस इरेक्टस (जावा मानव) जातीसारखीच आहे. या दोन्ही जातींची चालण्याची ढब ताठ उभी, शरीर सडपातळ चणीचे असून मस्तिष्क-गुहेचे आकारमान १,४०० घ.सेंमी. असलेल्या होमो निअँडरथलेन्सिस (निअँडरथल मानव) या जातीच्या मानाने
यांच्या मस्तिष्क-गुहेचे आकारमान कमी (९५० ते १,०७५ घ.सेमी.) आहे. जावा मानव अप्रगतिशील मानला जातो. जावातील होमो सोलोएन्सिस बहुधा जावा मानव व आधुनिक ऑस्ट्रॅलॉइड वंश यांच्यातील दुवा असावा.
निअँडरथलॉइड गट : हिमकाळातील शेवटच्या शीत टप्प्यातील पहिल्या अंतराहिमानीय काळातील निअँडरथल मानव बळकट चणीचा असून त्याच्या मस्तिष्क-गुहेचे आकारमान १,४०० घ.सेमी. होते. यूरोप, सहाराच्या दक्षिणेपर्यंतचा उत्तर आफ्रिकेचा भाग, पश्चिम आशिया इ. प्रदेशांतील पुष्कळ ठिकाणी कदाचित याचे म्हणता येतील असे काही परिवर्त आढळले आहेत. अल्जीरियातील दुसऱ्या अंतराहिमानीय टप्प्याच्या (मध्य प्लाइस्टोसीन) काळातील ॲटलँथ्रोपस हा होमो निअँडरथलेन्सिसचा परिवर्त असल्याचे दिसते व तो जावा मानवाला निअँडरथल मानवाशी जोडणारा दुवा असावा. जर्मनीमधील हिमकाळातील शेवटच्या शीत टप्प्यातील पहिल्या अंतराहिमानीय काळातील (ॲटलँथ्रोपसच्या काळातील) होमो स्टाइनहाइमेन्सिस जातीमधील आधुनिक मानवाच्या कवटीच्या वैशिष्ट्यांच्या जोडीने असलेल्या आदिम वैशिष्ट्यांवरून ही जाती जावा मानव व निअँडरथल मानव किंवा होमो इरेक्टस व होमो सेपिएन्स यांना जोडणारा दुवा मानली जाते. भरभक्कम जबडा व मागे झुकलेली हनुवटी असलेली होमो हायडेल्बर्गेन्सिस ही निअँडरथल मानवाची पूर्वगामी (पूर्वज) जाती आहे. ला चपेल-आक्स-सेंट्स येथे सापडलेले अवशेष हे निअँडरथल मानवाच्या अधिक मजबूत व रानटी मानवासारख्या परिवर्ताचे आहेत. प्लायोसीनच्या शेवटच्या काळात बहुधा होमो सेपिएन्स जातीद्वारे निअँडरथल गटाचा अंशतः लय झाला व त्याची जागा या जातीतील आधीच्या मानवांनी घेतली.
होमो ऱ्होडोसिएन्सिस गट : हा झँबियातील शेवटच्या अंतराहिमानीय काळातील गट आहे. यातील प्राण्यांच्या मस्तिष्क-गुहेचे आकारमान १,२५०-१,३२५ घ. सेंमी असून त्यांच्या कवटीची वैशिष्ट्ये होमो निअँडरथलेन्सिस जातीतील प्राण्यांच्या कवटीच्या वैशिष्ट्यांसारखी आहेत परंतु यांचे पाय पुष्कळसे होमो सोपिएन्ससारखे आहेत.
होमो सोपिएन्ससारखे तुल्य गट : केंट (इंग्लंड) येथील दुसऱ्या अंतराहिमानीय (पूर्व प्लाइस्टोसीन) कालीन ‘स्वान्स्कोंब मानव’ म्हणून ओळखले जाणारे अवशेष, केन्यातील होमा पर्वतात आढळले असून पूर्व प्लाइस्टोसीन कालीन कवट्यांचे व जबड्यांचे काही तुकडे आणि इझ्राएलमधील कॉर्मेल डोंगरात आढळलेले पहिल्या अंतराहिमानीय काळातील काही सांगाडे हे या गटात येतात. हे अवशेष बिनचूकपणे ओळखता .येत नाहीत. मात्र ते काहीसे होमो सोपिएन्ससारखे आहेत.
होमो सोपिएन्स : ग्रिमाल्डी (वायव्य इटली) येथे सापडलेल्या काही सांगाड्यांना ग्रिमाल्डी मानव असे संबोधण्यात येते आणि त्यांचा चेहरा वनात राहणाऱ्या आधुनिक आफ्रिकी मानवासारखा आहे. क्रो-मॅग्नॉन (दक्षिण फ्रान्स) भागातील एका गटाचे अवशेष उंच, पौर्वात्य प्रकारचे लंबकपाळ, मस्तिष्क-गुहेचे आकारमान १,६०० घ. सेंमी आणि हात व गुडघ्याखालील पाय लांब असलेले असे आहेत. दुसऱ्या गटातील प्राण्यांचे डोके पाश्चिमात्य प्रकारचे, रुंद असून त्यांना खरा क्रो-मॅग्नॉन मानव म्हणून संबोधतात व तो काही आधुनिक यूरोपियनांचा पूर्वज आहे. चान्सडेल (दक्षिण फ्रान्स) येथील अवशेषात एस्किमोसारखी वैशिष्ट्ये आढळतात आणि ते क्रो-मॅग्नॉन प्रकारचे परिवर्त असू शकतील.
पिल्टडाऊन मानव : इओअँथ्रोपस डाउसोनी म्हणजे पिल्टडाऊन मानव ही जाती कुतुहलजनक आहे. खालचा जबडा, एकत्र आणून जुळविलेले कवटीचे तुकडे व हातापायांची हाडे हे सर्व अवशेष पिल्टडाऊन (ससेक्स, इंग्लंड) जवळील नदीच्या वाळूत एकमेकांपासून अलग अलग असे पडलेले होते. ते गोळा करून त्यांच्या आधारावर ही जाती बनविण्यात आली आहे. खालचा जबडा निश्चितपणे कपीचा, बहुधा चिंपँझीचा, असल्याचे अलीकडेच दिसून आले आहे. हा जबडा कवटीशी जुळणारा नाही. कवटीची जाडी १२ मिमी. असून निअँडरथल मानव (जाडी सु.७ मिमी.) यांच्या कवट्यांपेक्षा ही कवटी बरीच जाड आहे या कवटीचे कपाळ उंच असून नाक सापेक्षतः बसके आहे. यामुळे हे कोडे सुटण्यासाठी आणखी काही सामग्री उपलब्ध होईपर्यंत थांबावे लागेल.
अशा प्रकारे जो काही पुरावा उपलब्ध आहे, तो अपूर्ण व विस्कळीत असा असला, तरी आधुनिक मानवापर्यंतच्या मुख्य टप्प्यांची मालिका पुढीलप्रमाणे असू शकेल झिंजांथ्रोपस-पिथे-कँथ्रोपस-ॲटलँथ्रोपस-होमो स्टाइनहाइमेन्सिस-होमो सोपिएन्स तुल्य.
दक्षिण आफ्रिकेतील ऑस्ट्रेलोपिथेकस या अपमानवावरून हा प्रदेश मानवाचे उत्पत्तिस्थान असल्याची एक मतप्रणाली आहे. मानव प्राण्याच्या जवळजवळ निश्चित अशा वाटणाऱ्या पूर्वजांबरोबर अवतरलेल्या शिवपिथेकसवरून पूर्व आफ्रिका मानवाचे उत्पत्तिस्थान असावे असा आणखी एक मतप्रवाह आहे. मानवाची ठळक वैशिष्ट्ये सर्वांत आधी भारतातील शिवालिक टेकड्यांत आढळलेल्या रामपिथेकसमध्ये आढळली असून त्यावरून भारत हा मानवाची जन्मभूमी असल्याचे सूचित होते. जावातील पिथेकँथ्रोपस इरेक्टसवरून व तेथेच आढळलेल्या आणि निअँडरथल मानवाशी जोडणारा दुवा म्हणून मानल्या गेलेल्या पिथेकँथ्रोपस सोलोएन्ससवरून जावा ही मानवाची ज्मभूमी असल्याचे काहींचे मत आहे.
तथापि मानवी वैशिष्ट्ये निश्चितपणे दाखविणारा सर्वांत पुरातन प्राणी रामपिथेकस हा होय आणि त्याच्याशी निकटचे संबंध असलेल्या मानवसदृश समूहाचा प्लायोसीन काळात शिवालिक प्रदेशात प्रत्यक्ष विकास होत होता म्हणून हा प्रदेश मानवाची जन्मभूमी असल्याचा पुरावा त्या भागात आढळण्याची दाट शक्यता आहे. अशा प्रकारच्या विविध दाव्यांमुळेच मानवाच्या जन्मभूमीविषयीचा निश्चित पुरावा उपलब्ध होईपर्यंत मानवाच्या जन्भूमीचा प्रश्न शिल्लक राहणार आहे.
भारतातील संशोधन कार्य : भारतासंबंधीच्या पुराप्राणिविज्ञानामध्ये आधीच्या काळात पडलेली भर ही मुख्यत्वे अंतरा-ट्रॅपी थरातील व तृतीय काळातील जीवाश्मासंबधीच्या माहितीची होती. हे काम जे. जी. माल्कमसन, एच्. जे. कार्टर, एस्. हिस्लॉप, आर्. हंटर, ई. फॉर्ब्झ यांसारख्या लष्करी अधिकाऱ्यांनी व धर्मगुरूंनी ईस्ट इंडिया कंपनीच्या कारकीर्दीत एकोणिसाव्या शतकाच्या मध्याच्या सुमारास केले. हे संशोधन कार्य स्फुट लेखांच्या स्वरुपात आहे व इंग्लंडमधील जिऑलॉजिकल सोसायटी ऑफ लंडन या संस्थेची पत्रिका (जर्नल) किंवा भारतातील एशियाटिक सोसायटी ऑफ बंगाल व एशियाटिक सोसायटीची मुंबई शाखा यांची पत्रिका वा सभेतील कामकाजाच्या नोंदी इत्यादींमध्ये हे लेख प्रसिद्ध होत असत. तथापि एच्. फॉक्नर (फाल्कनेर) व पी. टी. कॉट्ली यांनी शिवालिक टेकड्यांमधील पृष्ठवंशींच्या जीवाश्मांसंबंधी लिहिलेल्या फॉना अँटिक्वा शिवालेन्सिस (१८४५) हे पुस्तक असामान्य आहे.
इ. स. १८५१ साली ⇨ भारतीय भूवैज्ञानिक सर्वेक्षण संस्था स्थापन झाल्यावर भारतीय उपखंडातील जीवाश्म पद्धतशीरपणे गोळा करण्यासाठी योग्य ती परिस्थिती व यंत्रणा निर्माण होऊन प्रोत्साहन मिळू लागले आणि त्यामुळे अनेक संस्मरणीय अशा व्याप्तिलेखांना (प्रबंधिकांना) उपयुक्त अशी अतिशय मौल्यवान साधनसामग्री तज्ञांना उपलब्ध झाली. अशा तज्ञांपैकी अधिक महत्त्वाच्या व उल्लेखनीय तज्ञांची नावे व त्यांच्या संशोधनाचे विषय पुढे दिले आहेत. (१) एफ्. स्टोलिक्झ्का : दक्षिण भारतातील क्रिटेशस जीवाश्म (१८६३-७१) (२) ओ. फीस्टमँटल (फाइस्टमांटेल) : गोंडवनी वनस्पतिजीवाश्म (१८६५-८५) (३) डब्ल्यू. वागेन (१८७९-८७) व एफ्. आर्. कौपर-रीड (१९३१-४४) : (आता पाकिस्तानात असलेल्या) मिठाच्या डोंगरातील पुराजीव कालीन जीवाश्म (४) एम्. पी. डंकन व पी. स्लेडन (१८७१-८५), एम्. कॉस्मन व जी. पीसारो (१९०७-२७), तसेच ई. डब्ल्यू. व्रेडेनबर्ग (१९२६-२७): पश्चिम भारतातील (आता पाकिस्तानात) उत्तर क्रिटेशस व तृतीय कालीन जीवाश्म (५) एफ्. नोएटलिग : बलुचिस्तानातील (आता पाकिस्तानात) उत्तर क्रिटेशस कालीन जीवाश्म (१८९५-९७) (६) जे. डब्ल्यू. ग्रेगोरी (१८९३-१९००), एफ्. एल्. किचिन (१९००-०३), डब्ल्यू. वागेन (१८७३-७६), एल. एफ्. स्पॅथ (१९२७-३४), एल्. आर्. कॉक्स (१९४०-५२) : कच्छमधील जुरासीक कालीन प्राणिजात (७) डब्ल्यू. टी. ब्लॅनफोर्ड, टी. एच्. हक्सली, आर्. लिडेकर, एम्. जी. ईगरटन : तृतीय कालपूर्व पृष्ठवंशी (१८६५-८५) (८) आर्. लिडेकर व आर्. बी. फूट : तृतीय व तृतीयोत्तर कालीन पृष्ठवंशी (१८७४-८०) (९) सी. डीनर : हिमालयातील जीवाश्म (१८९५-१९०९) (१०) एफ्. आर्. कौपर-रीड : हिमालयातील व काश्मीरातील जीवाश्म (१९०८-३४) (११) जी. ई. पिलग्रिन : सस्तन प्राण्यांचे जीवाश्म (१९११-३४) (१२) बॅरन फोन ह्यूएन व सी. ए. मॅटले : क्रिटेशस कालीन डायनोसॉर (१९३३-४०). हे सर्व संशोधन कार्य भारतीय भूवैज्ञानिक सर्वेक्षण संस्थेच्या मेम्वार्स या रूपात पॅलिआँटालॉजिया इंडिकामध्ये आले आहे.
यानंतर भारतीय पुराप्राणिविज्ञानाच्या माहितीत पुढील तज्ञांनी भर घातली असून हे कार्य या बहुतेक सर्वजणांनी भारतीय भूवैज्ञानिक सर्वेक्षण संस्थेतील अधिकारी या नात्याने केले आहे: बी. प्रसाद, एस्. एल्. व्होरा (भारतीय प्राणिवैज्ञानिक सर्वेक्षण संस्था), डी. एन्. वाडिया, एम्. आर्. सहानी, एच्. एम्. लहरी, एन्. के. एन्. अय्यंगार, पी एन्. मुखर्जी, के जेकब, एम्. व्ही. ए. शास्त्री, बी. आर्. जे. राव, डी. एस. दासशर्मा, व्ही. डी. मामगैन, सी. त्रिपाठी, एस्. पी. सत्संगी, के एन्. प्रसाद.
भारतीय भूवैज्ञानिक सर्वेक्षण संस्थेच्या प्रकाशनांव्यतिरिक्त इतरत्र प्रसिद्ध झालेले भारतीय पुराप्राणिविज्ञानाविषयीचे उल्लेखनीय संशोधन कार्य पुढीलप्रमाणे आहे : (१) ए. डी. आर्किॲक व जे. हैम : पश्चिम भारतातील तृतीय कालीन जीवाश्म (१८५३) (२) एफ्. कॉस्माट : दक्षिण भारतातील उत्तर क्रिटेशस कालीन जीवाश्म (१८९५-९७) व डब्ल्यू. डी. मॅथ्यू (१९२९). ई. एच्. कोलबर्ट डब्ल्यू. के. ग्रेगोरी, एम्. हेल्मन व जी. के. ल्यूइस (१९३५) : शिवालिक प्राणिजात.
डेहाराडून येथे अलीकडेच स्थापन झालेल्या ऑइल अँड नॅचरल गॅस कमिशनमधील अनेक तज्ञांनी संशोधन करून अपतटीय व किनारी भागातील नवजीव व चतुर्थ कल्पातील गाळामध्ये असलेल्या सूक्ष्मजीवाश्मांसंबंधी नवीन माहिती मिळविली आहे. पणजी येथील नॅशनल इन्स्टिट्यूट ऑफ ओशनोग्राफी या संस्थेतील तज्ञ चतुर्थ कल्पातील सूक्ष्मजीवासंबंधी संशोधन करीत आहेत. भारतातील विद्यापीठांमधून पुराप्राणिविज्ञानाचे संशोधन कार्य १९२०-३० च्या सुमारास सुरू झाले. बंगलोर येथे एल्. राम राव यांनी सूक्ष्मपुराजीवविज्ञानाच्या संशोधनाचे केंद्र स्थापन केले. तेथे त्यांनी आपल्या सहकाऱ्यांच्या साहाय्याने संशोधन करून दक्षिण भारतातील क्रिटेशस व तृतीय कालीन फोरॅमिनीफेरा व रेडिओलॅरिया या प्राण्यांविषयीच्या माहितीत महत्त्वपूर्ण भर घातली आहे. बनारस विद्यापीठात राज नाथ यांनी गं. वा. चिपळूणकरांच्या साहाय्याने नर्मदा खोऱ्यातील बाघ थरांच्या पुराप्राणिविज्ञानविषयक संशोधनास सुरुवात केली. राज नाथ यांनी कच्छमधील जुरासिक कालीन जीवाश्मांसंबंधी सुरू केलेले संशोधनाचे कार्य तेथे पुढे चालू आहे. एम्. एस्. श्रीनिवासन यांनी अंदमान व निकोबार बेटांच्या सूक्ष्मपुराजीवविज्ञानावर लक्ष केंद्रित केले असून या बेटांवरील तृतीय काळातील थरांचे जैव-स्तरवैज्ञानिक दृष्टीने उपविभाग पाडण्यासाठी पाया घातला आहे. डी. के चक्रवर्ती यांनी प्रोबॉसिडिया गण व लॅमेटा थरांतील डायनोसॉर यांच्या जीवाश्मांचे अध्ययन केले आहे. लखनौ येथे एस्. आर्. नारायण राव व आर्. सी. मिश्रा यांनी गुजरात, राजस्थान व कच्छ या प्रदेशांतील सूक्ष्मजीवाश्मांच्या संशोधनाला प्रारंभ केला. शिवाय तेथील इतर शास्त्रज्ञ जैसेलमीर व कच्छ या भागाचा पुराजीवविज्ञानाच्या दृष्टीने अभ्यास करीत आहेत. चंडीगढ विद्यापीठात एम्. आर्. सहानी व आय्. खान यांनी शिवालिक शैलसमूहातील पृष्ठवंशी जीवाश्मांचे संशोधन सुरू केले असून तेथे कच्छमधील जुरासिक आणि काश्मीर व कुमाऊँ या प्रदेशांतील तृतीय कालीन जीवाश्मांचा अभ्यासही चालू आहे. हिमालयातील खडकांच्या थरातील कोनेडोंटविषयीचे व्ही. जे. गुप्ता यांचे कार्य विशेष उल्लेखनीय आहे. कारण भारतातील या गणाच्या जीवाश्मांवरील संशोधनास भारतात त्यांनी सुरुवात केली आहे. मद्रास विद्यापीठातील डी. ए. रशीद व अलीगढ येथील एस्. एन्. भल्ला हे दक्षिण भारतातील क्रिटेशस कालीन थर व आंध्र प्रदेशाच्या पूर्व किनाऱ्यावरील थर यांच्यातील सूक्ष्मजीवाश्मांचे संशोधन करीत आहेत. कलकत्याच्या इंडियन स्टॅटिस्टिकल इन्स्टिट्यूट या संस्थेतील टी. राय चौधरी व टी. एस्. कट्टी गोंडवन काळातील पृष्ठवंशी प्राण्यासंबंधी संशोधन करीत आहेत.खरगपूर येथे इंडियन इन्स्टिटयूट ऑफ टेक्नॉलॉजीतील ए. के. गुहा आणि बी. के घोष हे भारताच्या द्वीपकल्पातील क्रिटेशस कालीन खडकांतील ब्रायोझोआच्या जीवाश्मांचा अभ्यास करीत आहेत.
महाराष्ट्रात पुण्यामध्ये डेक्कन कॉलेज व महाराष्ट्र ॲसोसिएशन फॉर द कल्टिव्हेशन ऑफ सायन्स येथे पुराप्राणिविज्ञानविषयक संशोधन चालू आहे. डेक्कन कॉलेजात नर्मदा व गोदावरी नद्यांच्या खोऱ्यांतील चतुर्थ कल्पातील खडकांत आढळणाऱ्या पृष्ठवंशींच्या जीवाश्मांविषयी संशोधन चालू आहे. महाराष्ट्र ॲसोसिएशन फॉर द कल्टिव्हेशन ऑफ सायन्स या संस्थेत गं. वा. चिपळूणकर यांनी १९६५ साली भूविज्ञान व पुराप्राणिविज्ञान या विष्यांचा खास विभाग सुरु केला. तेथे त्यांच्या नेतृत्वाखाली नर्मदा खोऱ्यातील बाघ थर, तसेच त्रिचनापल्ली-पाँडिचेरी क्षेत्र व सौराष्ट्र येथील क्रिटेशस कालीन खडक यांच्यातील बृहत्जीवाश्मांसंबंधी सखोल संशोधन केले जात आहे. याशिवाय भारताच्या पश्चिम किनाऱ्यावरील चुतर्थ कल्पाच्या खडकातील सूक्ष्मजीवाश्म व कच्छमधील पुराजीवाविज्ञान याविषयीचे संशोधनही संस्थेने हाती घेतले आहे. मार्ग-चिन्ह जीवाश्मांचे पुष्कळ अध्ययन झाले असून अशा प्रकारचे हे भारतातील पूर्णतया नवे व पहिलेवहिले संशोधन कार्य आहे. बाघ थर, दक्षिण भारतातील क्रिटेशस कालीन व दक्षिण राजस्थानातील विंध्य संघातील खडक यांच्यात आढळणाऱ्या मार्ग-चिन्ह जीवाश्मांचे संशोधन येथे चालू आहे. गाळ साचण्याच्या वेळच्या परिस्थितीचे पुरापरिस्थितिविज्ञानाच्या दृष्टीने स्पष्टीकरण देण्याकरिता हे जीवाश्म विशेष उपयुक्त आहेत. पॉलिकीटा प्राण्यांच्या जीवाश्मांचे अध्ययनही भारतात येथेच प्रथम सुरू झाले असून बाघ थर व उत्तर क्रिटेशस कालीन त्रिचनापल्लीच्या प्रदेशातील थर यांच्यातील पॉलिकीटांच्या जीवाश्मांचे पुष्कळ अध्ययन येथे झाले आहे.
पहा : कँब्रियन; क्रमविकास; नवजीव; पुराजीव; पुराजीवविज्ञान; भूविज्ञान; मध्यजीव.
चिपळूणकर, गं. वा. (इं.); ठाकूर, अ. ना. (म.)
संदर्भ : 1. Beerbower, J. R. Search for the Past : An Introduction to Palaeontology, New Delhi, 1965.
2. Davis, A. M. An Introduction to Palaeontology, London, 1961.
3. Neaverson, E. Stratigraphical Palaeontology, London, 1962.
4. Romer, A.S. Vertebrate Palaeontology, Chicago, 1945.
5.Sahni, M. R. A. Century of Palaeontology Palaeobotany and Prehistory in India and Adjacent Countries, Lucknow, 1956.
6. Simpson G. G. Principles of Classification and Classification of Mammals, Bulletein of the American Museum of Natural History, Vol. 85, New York, 1945.
7. Wood, H. Palaeontology Invertebrate, London, 1961.
8. Zittel, Karl A. Von, Textbook of Palaeontology, 3. Vols., London, 1964.
९. ओक, शालिनी म. पुराजीवविज्ञान, पुणे १९७६.